




Составные компоненты молока, в том числе и ферменты, с момента доения и до получения готового молочного продукта неизбежно подвергаются разнообразным энергетическим воздействиям (механическим, тепловым, световым и др.), в зависимости от интенсивности и длительности которых изменяется их нативное состояние. Поглощаемая энергия ведет к изменениям тех или иных свойств ферментов и может служить в качестве энергии активации при разрыве химических или других связей или при образовании новых связей между составляющими молекулы фермента. Изменение третичной структуры белковой молекулы фермента под влиянием энергетических воздействий может привести к деформации активного центра и изменению каталитической активности фермента (гл. 1).
Получение молока. Основы качества молока закладываются на молочной ферме. В крупных молочных хозяйствах, где ручное доение коров заменено машинным с использованием высокопроизводительных доильных установок с центральным молокопроводом, молоко подвергается достаточно сильным механическим воздействиям. Степень этих воздействий зависит от конструктивных особенностей доильной установки (наличие резких подъемов молокопровода, применения молочных насосов, ввода молока из доильного аппарата в молокопровод перпендикулярно основному потоку молока, подсосу воздуха через доильный аппарат и неплотности соединений молокопровода), перепадов давления и температуры молока, при которых оно испытывает эти воздействия. В результате этого происходит прежде всего изменение дисперсности молочного жира, связанное с повреждением оболочек шариков жира (ОШЖ). При механическом воздействии на молоко вследствие возникающих усилий сдвига с ОШЖ удаляются микросомы и частично оболочечный белок. При столкновении шариков жира в первую очередь происходит взаимное проникновение их оболочек и возможное контактирование белковых слоев. При значительном механическом воздействии фосфолипидный слой становится проницаемым для жидкой фракции молочного жира, в результате чего происходит их агрегирование (см. рис. 15, с. 56). Следствием этих изменений является десорбция связанных с оболочкой шариков жира (ОШЖ) ферментов и, следовательно, повышение их активности в молоке.
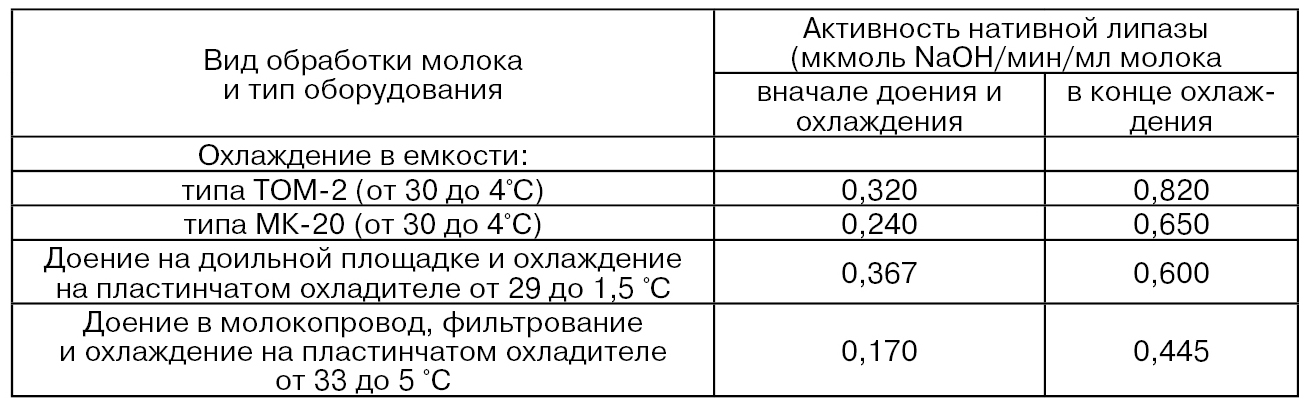
Исследования, проведенные Т. И. Безенко и И. П. Барановой, показывают, что при движении молока по линиям доильных установок УДТ-8 (протяженность молокопровода составляет 30 м с внутренним диаметром на различных его участках 15...36 мм) и АДМ-8 (протяженность молокопровода 60 м с внутренним диаметром 45 мм) приводит к увеличению содержания дестабилизированного жира соответственно на 125 и 209 % по сравнению с контролем, что составляет в абсолютном значении ≈ 3,5 и 5,4% к общему количеству жира в молоке. Механические воздействия, которые молоко испытывает в доильной установке с центральным молокопроводом, вызывают активирование липазы молока (табл. 32) (Шкинке В. Э.). Эти данные свидетельствуют о том, что повышение активности липазы молока в молокопроводе в зависимости от конструкции установок и особенностей их эксплуатации различно и колеблется от 0,11 до 0,25 мкмоль NaОН/мин/мл молока. При этом активность липазы в молокопроводе доильной установки повышается постепенно по мере продвижения молока.
Таблица 32
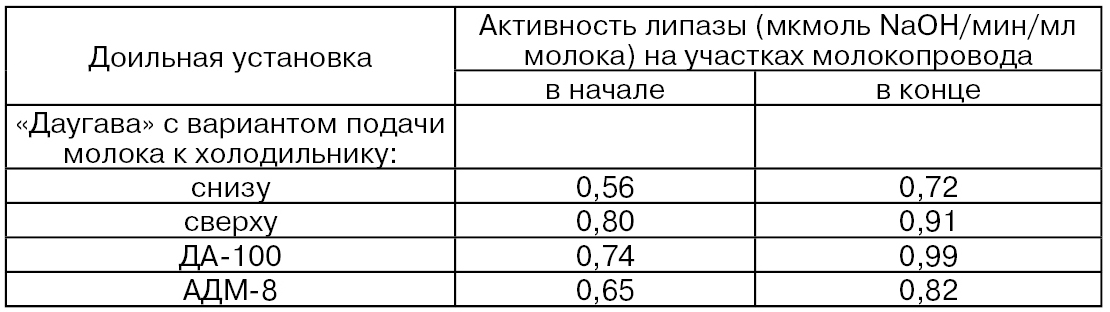
В результате повышения активности липазы и вследствие увеличенного содержания дестабилизованного жира в молоке повышается содержание СЖК. Так, при использовании установок УДТ-8 и АДМ-8 количество СЖК в исследуемых образцах молока, по сравнению с контрольным, увеличивалось от 37 до 42 % (Безенко Т. И., Баранова И. П.).
Исследования работы доильных установок различных типов показывают, что при крайне неблагоприятных в гидродинамическом отношении условиях монтирования трубопроводов для молока, малых диаметрах труб и высокой производительности насоса содержание СЖК в молоке вследствие активирования липазы может составить в среднем 1,56 мэкв./л по сравнению с 0,41 мэкв./л при доении коров машинным способом в переносные ведра, а максимально – соответственно 2,23 и 0,69 мэкв./л. Вместе с тем, содержание в молоке СЖК в количестве 2,0 мэкв./л и выше является пороговой величиной СЖК, влияющей на изменение вкуса молока (Тёпел А., 1979).
Отмечают следующую частоту возникновения липолиза в молоке, полученном при использовании различных способов доения: вручную – 12%, при машинном доении в переносные ведра – 14 и при доении в молокопровод – 33 %.
Ниже приведены данные по изменению степени липолиза в различных частях доильной установки с центральным молокопроводом при доении коров, находящихся на одной стадии лактации (по Fleming M.G, 1979):
|
Часть |
Липолитическая активность |
|
Доильный стакан |
0,88 |
|
Молокопровод: |
|
|
после 2 м и двух изгибов |
1,20 |
|
после 25 м |
і 1,43 |
|
Резервуар |
1,70 |
При этом в случае стойлового содержания липолитическая активность молока была выше, составляя пределы 0,93...1,85, чем в пастбищный период содержания коров – 0,76...1,04 СЖК мэкв./л.
Таким образом, в целях значительного снижения механических воздействий на молоко в доильных установках с центральным молокопроводом и, следовательно, предотвращения развития липолиза в молоке, приводящего к снижению его качества, необходимо строго соблюдать правила монтажа и эксплуатации доильных установок.
В зависимости от способа получения молока активность АТФ-азы в нем также различна. Так, в «цистернальном» молоке (полученное самотеком вне действия доильных рефлексов) активность фермента была почти в 2 раза ниже, чем в «рефлекторном» молоке (полученном из молочной железы при доении). Это обстоятельство очень важно учитывать для повышения АТФ-азной активности молока при использовании различных способов, вызывающих активную рефлекторную деятельность молочной железы при доении коров. Это связывают с соответствующей ролью АТФ-азы (подобно щелочной и кислой фосфатазам) в снабжении организма молодняка жизненноважным компонентом – неорганическим фосфором (М. П. Рязанский).
Машинное доение коров значительно облегчает труд, повышает его производительность, но при нарушении санитарно-гигиенических правил содержания оборудования, соприкасающегося с молоком, оно может оказаться очень серьезным источником первичного микробного обсеменения молока, а следовательно, и повышения его ферментативной активности.
Охлаждение и перемешивание молока. Содержание микроорганизмов в сыром молоке отражает уровень гигиены получения молока, особенно степень чистоты доильных установок, условия его фильтрации, охлаждения и транспортирования. Известны 2 пути обсеменения молока микроорганизмами: эндогенный – молоко обсеменяется непосредственно в молочной железе и экзогенный – обсеменение происходит из внешних источников – кожи животного, подстилочных материалов, кормов, воздуха, воды, доильной аппаратуры, рук и одежды работников молочной фермы. В железистой части молочной железы микроорганизмы могут находиться непостоянно и в единичном количестве КОЕ/мл. В выводных протоках и молочной цистерне количество бактерий может достигать уже десятков и сотен КОЕ/мл. В молоке (в сосковом канале), оставшемся от предыдущей дойки, постоянно размножаются бактерии, образуя бактериальную пробку, в которой количество бактерий достигает сотен тысяч КОЕ/мл молока. Молочная пленка, образующаяся в результате доения между кожей сосков и доильными стаканами, наличие на коже сосков различных складок, а также относительно высокая температура создают благоприятные условия для развития микрофлоры (Степаненко П. П., 1999).
В сыром охлажденном молоке имеется психрофильная микрофлора; она представлена в основном грамотрицательными аэробными бактериями родов Pseudomonas, Alcaligenes, Flavobacterium, Aeromonas, Achromobacter, количество которой к общему числу микроорганизмов, по данным различных исследователей, составляет: от 34 до 64 %; от 1 до 15 %; от 3 до 17 %; около 6 % и от 2 до 40 % соответственно. Среди превалирующих Pseudomonas наибольший % приходится на виды: Ps. fluorescens, Ps. fragi, Ps. viscosa, Ps. putrifaciens, Ps. graveolens (Law В. A., 1979).
Значимой по количеству групп психрофильной микрофлоры являются представители рода Acinetobacter, которые, как и Pseudomonas, в сырое молоко попадают обычно из воды. Для многих микроорганизмов способность проявлять психрофильные свойства принадлежит лишь отдельным штаммам, не являясь общевидовым признаком, например, некоторые представители плесеней, дрожжей, энтерококков. До 20 % штаммов Е. coli размножаются при 5...9 °С.
Наибольшее содержание психрофильных бактерий в сыром молоке отмечено в летний период – 1 · 106, осенью – 8 · 105, зимой и весной – 2,5 · 105 и 4 · 105 КОЕ/мл соответственно. При переводе коров со стойлового содержания на пастбищное, количество психрофильных бактерий в молоке заметно увеличивается. При стойловом содержании коров в молоке преобладают бактерии рода Acthrobacter, а при пастбищном – наиболее часто обнаруживаются Flavobacterium, Alcaligenes и Acthrobacter.
В сыром охлажденном молоке обнаружены и грамположительные бактерии, но в значительно меньших количествах, чем грамотрицательные. Они представлены в основном родами Bacillus, Acthrobacter, Micrococcus.
Мезофильная микрофлора сырого молока представлена в основном бактериями родов лактококков и микрококков, минимальная температура роста, например, молочнокислых бактерий составляет 10 °С. В сыром молоке постоянно присутствуют бактерии группы кишечной палочки (по Моисеевой Е. Л., 1988).
В молоке, полученном в хороших санитарных условиях, преобладает мезофильная микрофлора – лактококки и микрококки, а при неудовлетворительных санитарных условиях – психрофильные бактерии рода Pseudomonas (лактококки составляли 10...20 % от общего количества бактерий, а аэробные спорообразующие бактерии – до 12 %). Показано, что в молоке, охлажденном непосредственно после доения коров, содержалось от 7 · 103 до 3 · 104 КОЕ/мл психрофильных бактерий. Если в молоке, полученном в хороших санитарных условиях, содержалось около 10% психрофильных бактерий от общего количества микрофлоры, то в молоке, полученном в неудовлетворительных санитарных условиях – до 75 % (Моисеева Е. Л., 1988).
В странах ЕС и США принятая норма содержания микроорганизмов в молоке-сырье составляет не более 105 КОЕ/мл. Те же показатели для молока высшего сорта рекомендует и ММФ. В соответствии с ГОСТ Р 52054, норма содержания микроорганизмов в молоке высшего сорта – не более 3 · 105 КОЕ/мл, для сортового молока в целом – не более 4 · 106 КОЕ/мл.
С целью предотвращения или снижения возможного продуцирования микрофлорой молока-сырья различных экзоферментов, проявление активности которых может вызывать снижение качества молока при его хранении, необходимо как можно скорее охладить молоко, при этом от скорости охлаждения молока после доения и в дальнейшем – температуры и длительности его хранения будет во многом зависеть микробиологическое качество молока-сырья и его физико-химические показатели.
Ниже приведены данные по влиянию процесса доения и технологии первичной обработки молока на его бактериальную обсемененность (Улитенко А. И., Соколовский Э. И., Пушкин В. А., 2005):
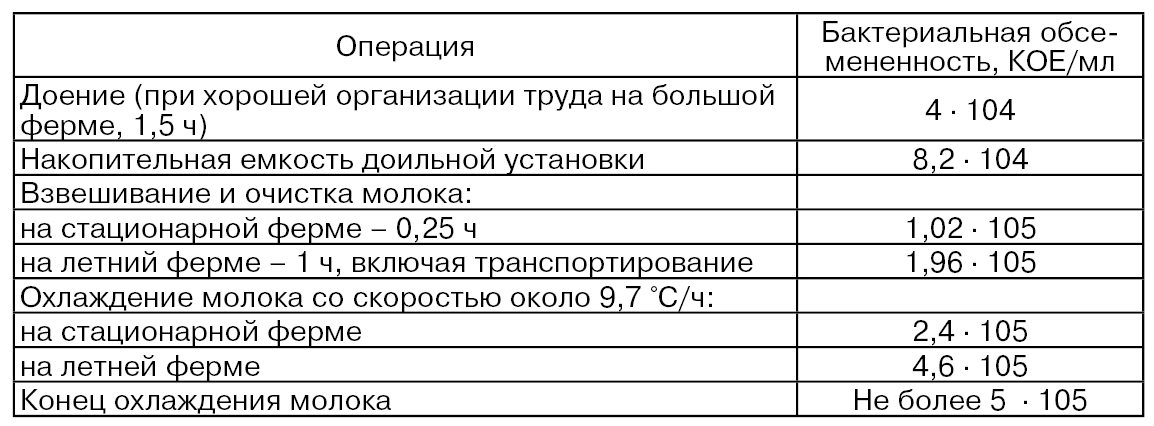
Использование на практике тех или иных режимов охлаждения и перемешивания является наиболее важными факторами первичной обработки молока, влияющими на изменение количества и свойств его ферментов.
Парное молоко имеет температуру 37 °С, и все составные компоненты его находятся в нативном состоянии. Понижение температуры способствует некоторому изменению состояния компонентов молока. При этом, прежде всего, затрагиваются гидрофобные связи в ОШЖ и мицеллах казеина, которые при низких температурах не так устойчивы, как при повышенных.
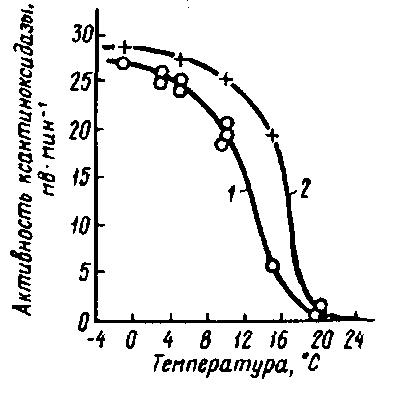
Рис. 31. Изменение активности ксантиноксидазы сырого молока при охлаждении (Back W. D., et al.):
1 – 2,22% жира, 3,04% белка; 2 – 3,24% жира, 3,04% белка
Одним из последствий неустойчивости гидрофобных связей является десорбция ксантиноксидазы из оболочек шариков жира молока, что ведет к значительному повышению ее активности в водной фазе (рис. 31). При этом отмечается прямая зависимость активности фермента от содержания в молоке жира. Для десорбции имеет значение скорость охлаждения молока. При низкой скорости охлаждения (1,5°С/ч), а следовательно, и более продолжительном (около 20 ч) снижении температуры молока, например с 37 до 6°С, имеет место заметная десорбция ксантиноксидазы, в то время как при высокой скорости охлаждения молока (8°С/ч) и соответственной продолжительности (2...4 ч) его охлаждения десорбция этого фермента незначительна. Кроме того, охлаждение почти всегда связано с механической нагрузкой на молоко. Поэтому повышение скорости движения потока охлажденного до 6°С сырого цельного и обезжиренного (0,4% жира) молока с 1...2 м/с (ламинарное движение) до 4...6 м/с (турбулентное движение) приводит к заметной десорбции ксантиноксидазы с оболочки шариков жира (ОШЖ) молока. Активность фермента по сравнению с начальной активностью возрастает соответственно почти на 60 и 260 % при 30-минутной продолжительности такой нагрузки и на 20 и 80 % при нагрузке в течение 15 мин (рис. 32). При этом количество фермента, имеющегося в молоке, не увеличивается, а происходит только его активация. Силы, действующие на ОШЖ, не только способствуют выделению фермента из ОШЖ, но и вызывают расщепление его на субъединицы, что повышает активность имеющейся в молоке ксантиноксидазы (Back W. D., Reuter Н., Rotter Z., Jakubowski M., 1981).
В связи с тем, что 20...30% нативной каталазы связано с оболочками шариков жира молока, то при охлаждении, видимо, может произойти некоторая десорбция этого фермента в водную фазу молока, а также расщепление молекулы фермента на субъединицы. Следовательно, активность каталазы в молоке может повыситься.
Перемешивание является наиболее распространенной формой механического воздействия на молоко при его транспортировке по трубопроводам, заполнении и опорожнении емкостей и т. д. При этом существенное значение имеет температура молока. При интенсивном перемешивании охлажденного молока путем аэрации в результате происшедшего ранее ослабления гидрофобных связей при его охлаждении, а также частичного изменения структуры ОШЖ наряду с ксантиноксидазой и частично с каталазой происходит десорбция щелочной и кислой фосфатаз. Например, при перемешивании охлажденного цельного молока (10 °С) струей воздуха, азота или кислорода со скоростью 20 мл/мин в течение 20 мин более 95% общей активности щелочной фосфатазы (как и ксантиноксидазы) было обнаружено в обезжиренной фракции молока. В связи с этим считают, что заметное повышение активности вышеуказанных ферментов в обезжиренной фракции молока может служить тестом степени подсбивания жира в молоке (Bhavadasan M. K., Abrahum M. J., Ganduli N. C., 1982, McPherson A.V., Kitchen B.J., 1983).
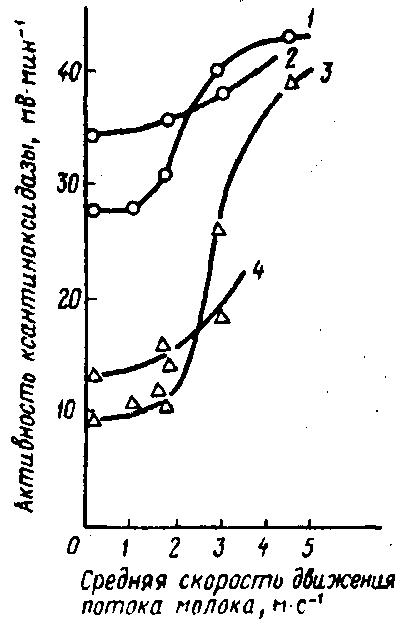
Рис. 32. Активность ксантиноксидазы в цельном и обезжиренном сыром молоке в зависимости от скорости течения потока молока:
1 – цельное молоко при продолжительности нагрузки 30 мин; 2 – цельное молоко при продолжительности нагрузки 15 мин; 3 – обезжиренное молоко при продолжительности нагрузки 30 мин; 4 – обезжиренное молоко при продолжительности нагрузки 15 мин
Следствием ослабления гидрофобных связей в мицеллах казеина при охлаждении молока и механическом воздействии перемешивания на молоко является и повышение в нем активности нативных липаз, о чем свидетельствуют данные, приведенные в табл. 33. Если при охлаждении сырого молока в емкостях ТОМ-2 и МК-20, сопровождающемся его перемешиванием мешалками, повышение активности липаз составило 155 и 170% соответственно, то при охлаждении молока на пластинчатом охладителе – только 63 % первоначальной активности. В молокопроводе доильной установки вследствие взбалтывания и вспенивания молока активность липазы повышалась на 162% первоначальной активности (Олконен А. Г.).
Таблица 33
Результаты исследований показывают, что при скорости турбулентного движения молока более 6 м/с наблюдается активация липазы, а следовательно, и повышение содержания СЖК (почти в 2 раза по сравнению с контролем). Эти изменения усиливаются с повышением температуры (от 5 до 25 °С) и продолжительности процесса (от 7 до 15 мин) (Rotter Z., Jakubowski M., 1981).
Низкие скорости движения потока молока в молокопроводах также приводят к активации липазы, так как при небольшом количестве молока, смешанном с большим количеством воздуха, создается пульсирующее движение в системе. Такое явление может встречаться там, где доение коров осуществляется медленно, доильные стаканы на сосках молочной железы остаются после окончания доения, а также если применяют вакуумные насосы недостаточной мощности.
Степень воздействия механического перемешивания на активность ферментов молока зависит, например, от вида используемых мешалок. В табл. 34 приведены данные по изменению содержания СЖК при перемешивании молока в охлаждаемых ваннах и резервуаре для хранения молока. Установлено, что при перемешивании молока в резервуарах меньший вихревой поток отмечается в его середине. Степень липолиза более выражена на поверхности молока, что обусловлено пенообразованием. Увеличение продолжительности и интенсивности перемешивания повышает выраженность липолиза в 1,5...2,0 раза (Kirst Е., 1980).
Таблица 34
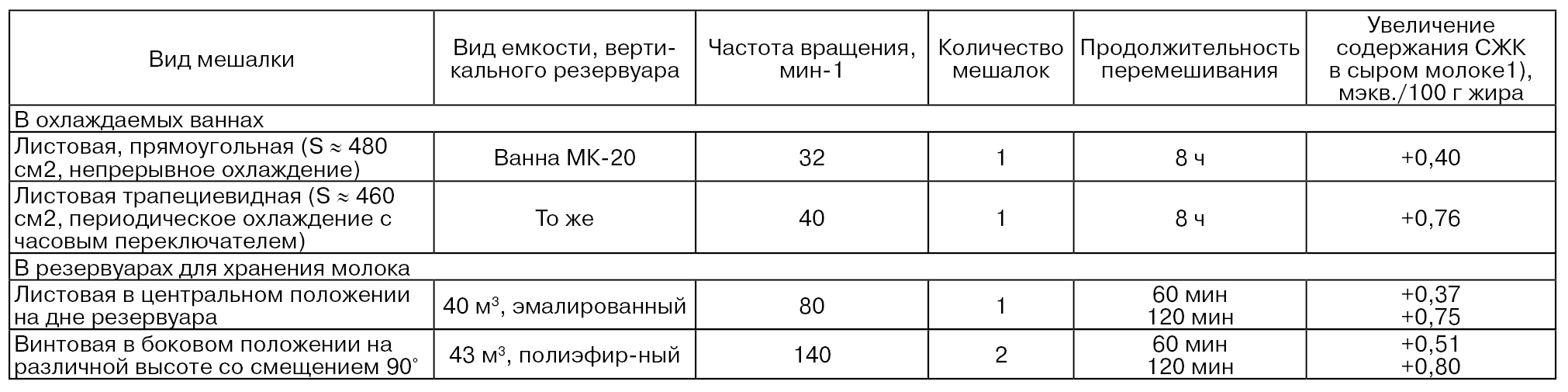
1) При перемешивании, по сравнению с исходным содержанием.
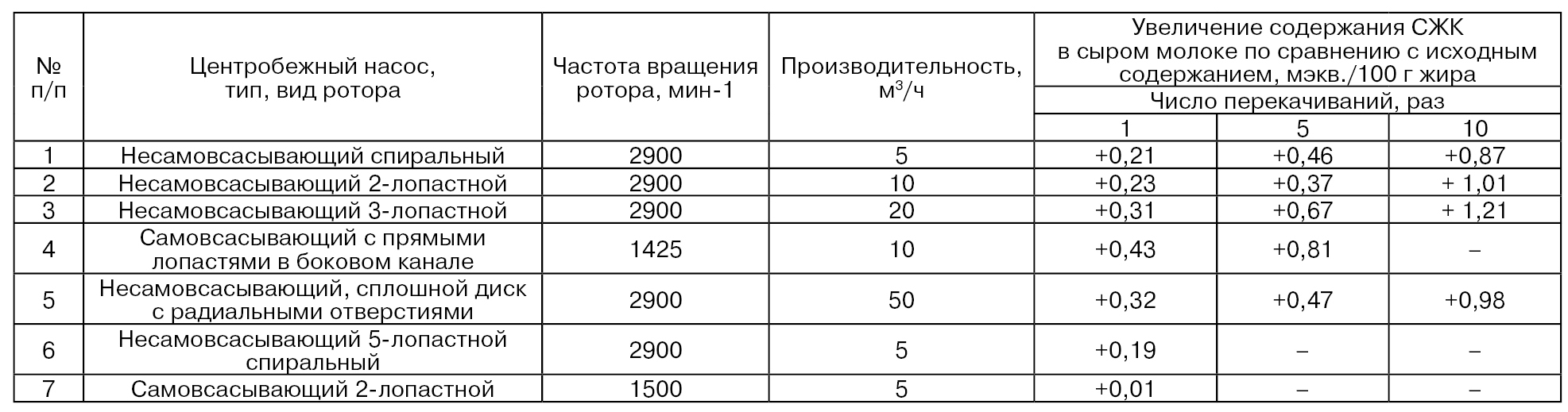
Степень механического воздействия на молоко зависит от вида используемых насосов (табл. 35). Приведенные данные показывают, что в насосах 6 и 7 механическое воздействие на молочный жир было незначительным. При работе насосов 3, 4 и 5 наблюдались относительно сильные изменения молочного жира. Таким образом, на практике следует отдавать предпочтение тем насосам, при использовании которых происходит наименьший липолиз молока (Kirst Е., 1980).
Основной причиной индуцированного липолиза является интенсивное смешивание молока с воздухом (воздух частично дестабилизирует оболочку шарика жира). Это происходит при прохождении молока через молокопроводы или вследствие любой обработки, вызывающей сильное взбалтывание или встряхивание. В молоке, перемешанном при отсутствии воздуха, индуцированный липолиз проявляется гораздо в меньшей степени (характер воздействия других газов, например азота, такой же, как и воздуха).
Таблица 35
Считают, что вспенивание усиливает активность липазы за счет увеличения числа шариков жира с нарушенной структурой оболочки и адсорбции на них казеина, с которым в основном связаны липазы молока. Для снижения степени индуцированного липолиза важно, чтобы во время механического воздействия на молоко его температура была как можно ниже. Холодное свежее молоко (5 °С) более устойчиво к механическим воздействиям перемешивания со вспениванием, чем теплое молоко, однако после определенной продолжительности его хранения в результате нарушения структуры ОШЖ оно более подвержено такому влиянию.
Липолиз, вызванный перемешиванием, быстро распространяется, но это происходит лишь в течение сравнительного короткого периода времени. Дальнейшего накопления СЖК не наблюдается, несмотря на избыточное количество субстрата. Например, липолиз, индуцированный барботированием газа через теплое молоко (30 л/мин), что вызывает турбулентность и пенообразование, выравнивается через 10...15 мин, когда количество СЖК приближается к 3,5 мэкв./л. Выравнивание индуцированного липолиза в молоке с течением времени объясняется накоплением СЖК на поверхности раздела фаз жир/вода, которые являются ингибиторами липаз, а также неспособностью ферментов десорбировать с этой поверхности раздела. В некоторых случаях этот процесс обусловливается синтезом липидов из образующихся жирных кислот. Все это приводит к замедлению и в конечном итоге к подавлению липолиза, если перемешивание молока не повторяется (Downey W. К., 1980).
Активирование индуцированного липолиза может происходить и в результате резкого изменения температуры молока. Если свежее сырое молоко, которое было охлаждено до 5 °С и ниже, нагреть до 25...35 °С, затем охладить и оставить на хранение, то может произойти липолиз. Это обусловлено изменением структуры поверхности оболочек шариков жира и, следовательно, большим контактом липазы с молочным жиром.
Влияние охлаждения молока как единственного фактора «немолочного» происхождения на возникновение спонтанного липолиза молока связано с аглютинированием шариков жира иммуноглобулинами, с которыми частично связаны нативные липолитические ферменты.
Представляет интерес рассмотрение вопроса и об изменении протеолитической активности молока при его охлаждении. Как было показано, протеиназы молока в основном связаны с мицеллами казеина, а именно с β-казеином, с которым они находятся в гидрофобной связи (рис. 14, а). При температуре ниже 10 °С вследствие заметного ослабления гидрофобных взаимодействий в мицелле казеина она частично распадается на субмицеллы, и β-казеин (основная гидрофобная фракция) выходит из мицеллы и в дальнейшем подвергается гидролизу при хранении молока. Следовательно, при охлаждении молока возможность возникновения протеолиза в нем усиливается (Reimendes Е. Н., Herlitz Е., 1979).
Наряду с рассмотренными ферментами, активность которых заметно изменяется при охлаждении молока вследствие их связи с ОШЖ или мицеллами казеина, активность многих других ферментов, связанных с указанными компонентами молока, также может измениться, что повлечет за собой соответствующие изменения качества
сырого молока (гл. I).
Хранение и транспортирование молока. Время, в течение которого микроорганизмы в свежевыдоенном молоке не развиваются и даже частично отмирают, составляет бактерицидную фазу, которая обусловлена присутствием в молоке нативных компонентов, обладающих бактериостатическим и бактерицидным действием. Продолжительность бактерицидной фазы зависит от температуры хранения молока, степени его исходного микробного обсеменения и состава микрофлоры, а также индивидуальных особенностей животных. Чем выше температура хранения молока, тем короче бактерицидная фаза, что видно из нижеприведенного:
|
Температура хранения молока, °С |
37 |
30 |
25 |
10 |
5 |
2 |
|
Продолжительность бактерицидной фазы, ч |
2 |
3 |
5 |
12...20 |
24...30 |
До 48 |
По окончании бактерицидной фазы начинается размножение микроорганизмов, интенсивность которого зависит от их вида. Этот период продолжается от 12 ч до 1...2 сут. При этом микрофлора молока может заметно увеличиться от нескольких тысяч, которое оно имеет к концу бактерицидной фазы.
В зависимости от температуры хранения в этот период в молоке может развиваться микрофлора низких и средних температур. Микрофлора низких температур развивается при хранении молока от 0 до 10 °С, причем это развитие идет очень медленно. Например, при температурах 4...5 °С накопление биомассы за 24 ч составляет 9 %. При этом молочнокислые бактерии практически не размножаются. На основании многих литературных данных, в сыром молоке, охлажденном до температуры 3...5 °С, за 2 сут хранения количество психрофильных микроорганизмов может увеличиться в среднем на порядок, что напрямую зависит от исходного уровня бакобсемененности молока. В хранении молока при низких температурах психрофилы становятся доминирующей микрофлорой. Способность психрофилов размножаться при низких температурах связывают с особенностями ферментов и липидов в цитоплазматической мембране, а именно в липидах содержится повышенное количество ненасыщенных жирных кислот, вследствие чего мембрана постоянно находится в жидкокристаллическом состоянии. При низких температурах у психрофилов наблюдается увеличение образования экзоферментов, с помощью которых происходит более активное расщепление различных субстратов и поступление их составных частей в клетку. Микрофлора средних температур развивается при хранении молока от 10 до 35 °С. При этом характерно быстрое размножение и неуклонное нарастание количества молочнокислой микрофлоры, которая доминирует заметно уже над остальными микроорганизмами. Однако в составе микрофлоры, особенно в начальной стадии фазы смешанной микрофлоры, развиваются бактерии группы кишечной палочки, флюоресцирующие и другие гнилостные бактерии (Королева Н. С., Семенихина В. Ф., 1980; Степаненко П. П., 1999; Свириденко Г. М., Захарова М. Б., 2005).
В условиях холодильного хранения ферментативная активность молока в той или иной степени повышается. Это происходит как в результате воздействия режимов охлаждения на отдельные компоненты молока и как следствие – на активность нативных ферментов молока, так и в результате жизнедеятельности микроорганизмов, особенно при нарушении санитарно-гигиенических условий получения, первичной обработки и транспортировании молока. В молоке обнаружена активность около 60 ферментов микробного происхождения, которые наряду с ферментами нативного происхождения катализируют различные процессы, вызывающие изменения компонентов молока и ухудшающие его качество при хранении.
Изучение изменения микрофлоры сырого молока при его хранении показало, что в сборном молоке, полученном с фермы, общее количество аэробных мезофильных бактерий в течение первых суток его хранения при 13...15°С увеличилось в 30 раз, при 8...10°С примерно в 20 раз, а при 3...5 °С – только в 3 раза. При этом количество бактерий группы кишечной палочки увеличилось соответственно в 13, 8 и 2 раза, психротрофильных бактерий – в 14, 3,5 и 2,5 раза и холодостойких молочнокислых бактерий – соответственно в 30, 10 и 2 раза (Королева Н. С, Семенихина В. Ф., 1980).
Своевременное, эффективное охлаждение молока и быстрая доставка его на предприятия промышленности (не более 12 ч после доения коров) позволяют сохранить исходную редуктазную активность молока. Глубокое охлаждение молока (ниже 5 °С) в целях его длительного хранения влечет за собой изменение качественного состава микрофлоры и ее биохимической активности. Преобладающей становится психрофильная микрофлора. При этом связь между общей редуктазной активностью молока и общим числом в нем микроорганизмов становится менее выраженной. В то же время отмечается довольно четкая зависимость между микробным обсеменением такого молока и содержанием в нем пировиноградной кислоты как основного продукта метаболизма микроорганизмов (особенно грамотрицательных). Наряду с пировиноградной кислотой при развитии в молоке микроорганизмов образуются различные соединения, сообщающие молоку несвойственные ему пороки вкуса и запаха (рис. 33). При этом накопление пирувата (и лактата) идет постепенно в течение всего процесса первичной обработки (включая охлаждение и хранение) молока (табл. 36) (Suhren G., 1982, 1993).
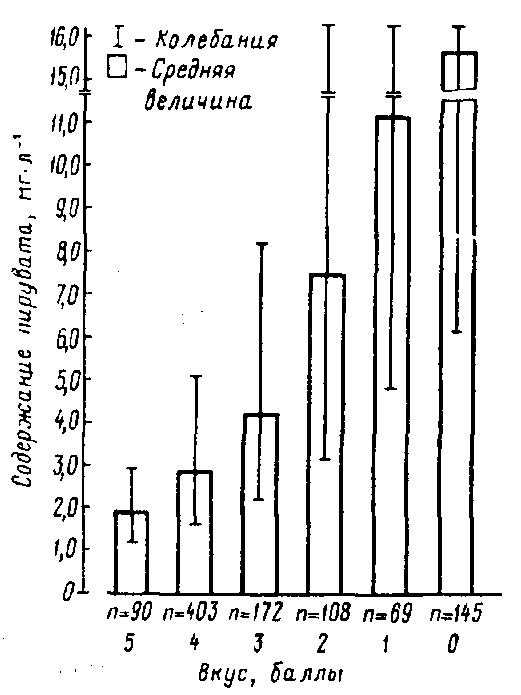
Рис. 33. Соотношение между оценкой вкуса и содержанием пирувата в сыром молоке (n – количество образцов молока) (Suhren G., 1993)
Установлена обратная зависимость между содержанием в сыром молоке пирувата и его сохраняемостью.
Следует отметить, что психротрофильные микроорганизмы при хранении сырого охлажденного молока благодаря их гетероферментативной природе могут явиться причиной возникновения окисленного привкуса, не говоря о том, что вследствие высокой протеолитической активности эти микроорганизмы являются хорошим источником азота, необходимого для жизнедеятельности мезофильных молочнокислых бактерий – активных кислотообразователей молока.
В связи с тем, что активность ксантиноксидазы в молоке связана с возникновением в нем окисленного привкуса, представляет интерес рассмотреть вопрос о влиянии на эту активность режимов хранения молока. Результаты работ некоторых исследователей, приведенные в табл. 37, показывают, что активность ксантиноксидазы в условиях хранения молока при 2...5 °С повышается в основном в первые 12...24 ч, а затем остается на одном уровне или несколько снижается (Cerbulis J., Farrell Н. М., 1977; Bhavadasan М. К., Ganguli N. С., 1980).
Таблица 36
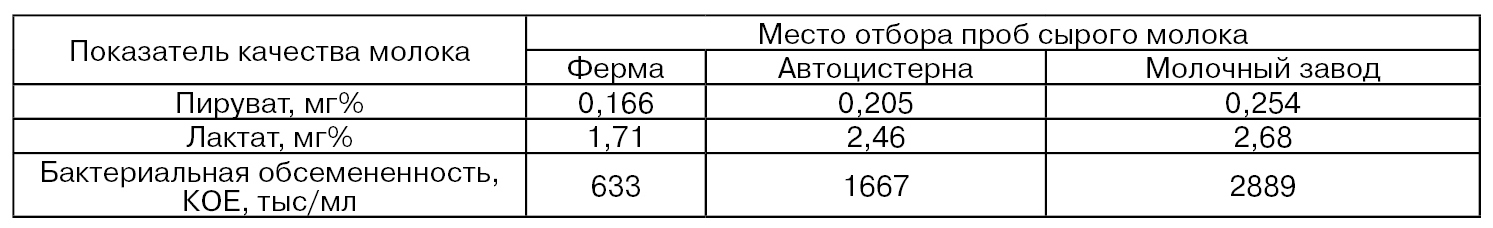
Таблица 37
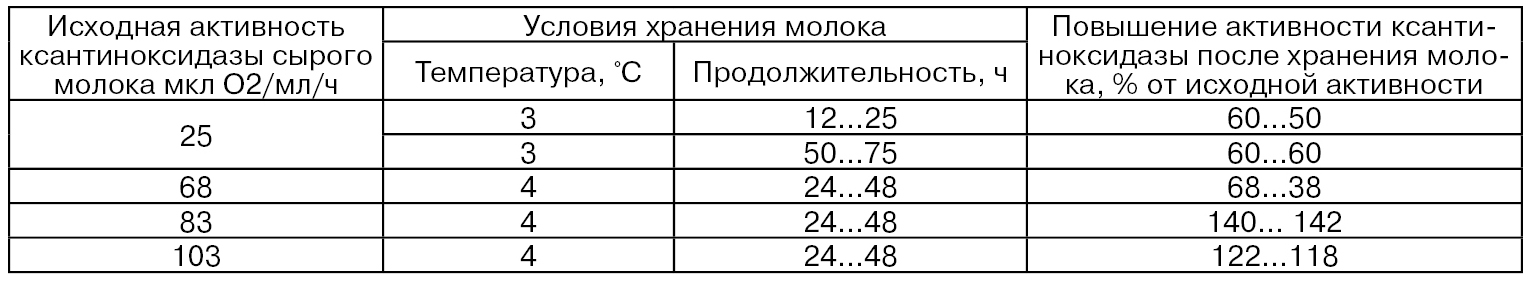
Одной из причин повышения активности ксантиноксидазы при непродолжительном хранении молока является переход ее D-формы в О-форму. Так, в процессе хранения молока в течение 16 ч при 4°С активность ксантиноксидазы возрастает от 52 до 75 ед., а ксантиндегидрогеназы – снижается с 17 до 10 ед. (Nakamura М., Yamazaki J., 1982).
При охлаждении и последующем хранении сырого молока происходит некоторое снижение количества оболочечного материала шариков жира, например, с 1280 мг на 100 г жира до 910 мг на 100 г жира. Вследствие этого происходит некоторое перераспределение активности ксантиноксидазы между жировой и водной фазами молока (табл. 38) в сторону увеличения активности в последней.
Причиной некоторого снижения активности ксантиноксидазы при длительном хранении молока может явиться и частичный гидролиз этого фермента протеазами молока (Zikakis J. P., Silver M. R., 1984).
При хранении сырого молока увеличивается и активность каталазы, что связано в основном с жизнедеятельностью микроорганизмов.
Таблица 38
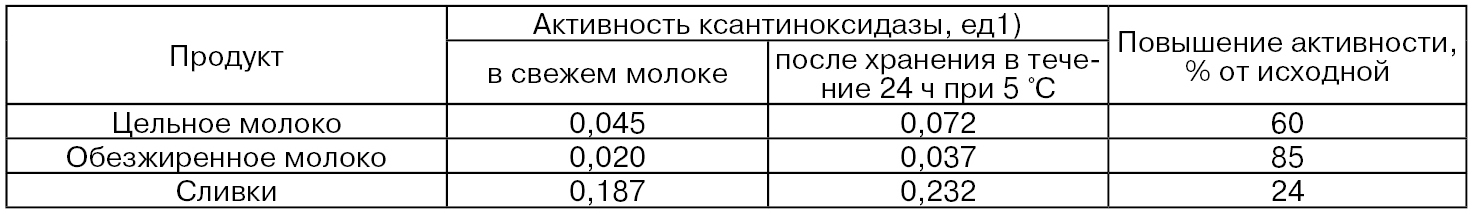
1) 1 ед. соответствует количеству фермента, вызывающего изменение поглощения на 1,0/мин при 290 нм.
Так, показано, что активность каталазы в сыром свежевыдоенном молоке (с трех ферм) в весенне-летний период составляла 8...17 и в осенний период – 20...25 % кислорода. В условиях хранения этого молока при 3...5°С в течение 2 сут в весенне-летний период активность каталазы значительно не изменялась, при 8...10°С хранения в течение суток она повышалась на 2...10 %, а в течение вторых суток она продолжала увеличиваться, но не выходила за пределы 20 % кислорода. В осенний период года активность фермента в молоке в первые сутки хранения при 3...5°С повышалась более заметно, чем в весенне-летний период, при 8...10°С к концу первых суток хранения она заметно увеличивалась и в течение следующих суток составила 30...35 % кислорода (Шидловская В. П., Патратий А. П., Насонова Л. М., 1985).
При хранении молока несколько изменяется активность лактопероксидазы. Так, если в процессе хранения при 3 °С в течение 2 сут активность фермента в молоке практически не изменяется, то через 4 сут его активность увеличилась на 30 %. Считают, что это происходит в результате снижения количества связанной лактопероксидазы и перехода ее в свободное, более активное состояние, а также вследствие расщепления фермента на субъединицы (что характерно и для каталазы и ксантиноксидазы).
В ряде стран изучена возможность повышения стойкости в хранении охлажденного и неохлажденного молока за счет активирования лактопероксидазо-антибактериальной системы молока, состоящей из трех компонентов: самого фермента – лактопероксидазы, тиоцианата и перекиси водорода[1]. Показано, что для проявления активности этой системы необходимо наличие в молоке лактопероксидазы в количестве 1...2 мкг/мл или по активности больше или равной 0,17 нкат, что значительно меньше таковых величин, характерных для сырого молока (Björck L., 1982; Griffits М. W., 1986; Zarung S. A., Gnan S. О., 1999; Villamiel M., Jong P., 2000). Естественное содержание тиоцианата в коровьем молоке составляет 0,1...1,5 мг%, однако чаще оно равно 0,1...0,5 мг% (в молоке других сельскохозяйственных животных 0,6...0,8 мг%) (Abb. El. Ghani S., Sayed R. F., 1997), что достаточно для проявления лактопероксидазо-антибактериальной системы. Для активации этой системы рекомендуется повысить естественную концентрацию тиоцианата до 1...1,5 мг%.
Результаты анализа 120 образцов молозива и молока в первые 30 дней лактации коров показали, что содержание тиоцианата постепенно снижалось с 1,26 до 0,53 мг/% (Uysal Н. R., 1996). Следует отметить, что результат возможного проявления в молоке активности роданазы может быть дополнительным источником содержаниния в молоке тиоцианата. Перекись водорода в бактериально чистом молоке практически не обнаруживается. В аэробных условиях многие лактобациллы, лактококки и стрептококки могут продуцировать некоторое количество перекиси водорода. Ее источником могут быть реакции, катализируемые аминоксидазой, глюкозоксидазой, ксантиноксидазой, сульфгидрилоксидазой и пероксид-дисмутазой (гл. 1). В связи с этим возникает необходимость внесения в молоко дополнительного количества перекиси водорода. Установлено, что для проявления заметного антибактериального эффекта молока достаточно внести в молоко перекись водорода в количестве 0,85 мг%.
Активность лактоперексидазо-антибактериальной системы замедляет рост грамотрицательных бактерий (например, сальмонеллы, колиформы), но может вызывать и их гибель, в то время как у грамположительных бактерий (например, стрептококки, лактобациллы) она ингибирует их рост, но не вызывает их гибель. Такую разную чувствительность бактерий к действию лактоперексидазной системы можно объяснить различием в строении их клеточной мембраны и ее пропускной способности. Действие этой системы состоит в окислении тиоцианата с образованием, в частности, промежуточных продуктов – цианосерной/цианосернистой кислот и гипотиоцианата:
![]()
Гипотиоцианат, как сильный окислитель, воздействует на клеточную мембрану микроорганизмов, вызывая подавление синтеза белков, РНК, ДНК, в результате чего ингибируется их жизнедеятельность или же происходит лизис клеток.
Действие лактопероксидазо-антибактериальной системы на микрофлору молока более специфично, чем действие чистой перекиси водорода, которую добавляют в молоко для достижения антибактериального эффекта в количестве 30...80 мг%. Такие высокие концентрации перекиси водорода инактивируют лактопероксидазу молока, в то время как добавление перекиси водорода в количестве 1...1,5 мг% немедленно и полностью используются в ферментативной реакции.
Некоторые результаты исследований по активированию лактопероксидазо-антибактериальной системы сырого коровьего молока
и молока других сельскохозяйственных животных приведены в табл. 39.
Таблица 39
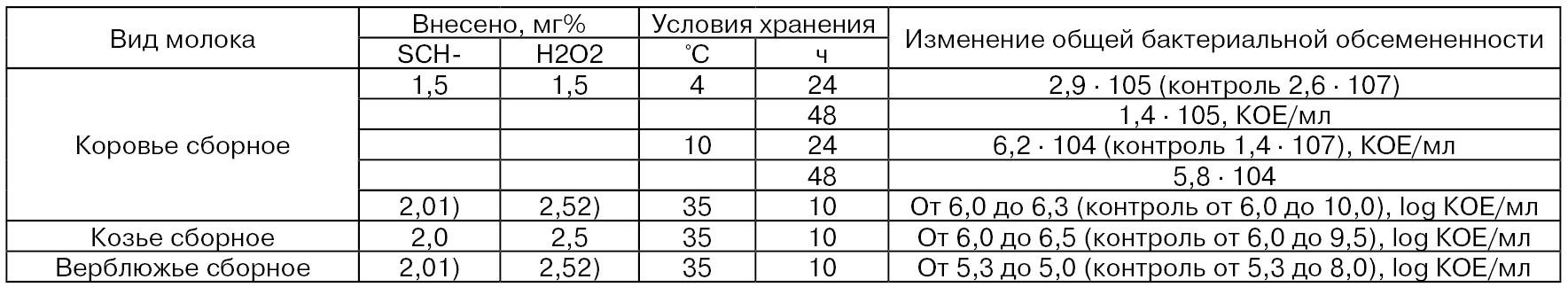
1) Количество внесенного тиоцианата натрия – NaSCN.
2) Количество внесенного перкарбоната натрия – 2Na2CO3 • 3Н2O2.
Потребление молока, в котором активирована лактопероксидаза, не представляет опасности для здоровья человека, поскольку продукты окисления тиоцианата нестойки и разрушаются при тепловой обработке.
При использовании тиоцианата и Н2О2 для активирования лактопероксидазы молока, Н2О2 утилизируется полностью, тогда как остаточные количества тиоцианата к концу хранения молока могут составить от 15 до 75 % (в зависимости от условий хранения и вида молока).
Вместе с тем, остающееся возможное количество тиоцианата — от 0,3 до 1,2 мг% небольшое по сравнению с естественной концентрацией этого вещества в слюне человека (5...30 мг%) и желудочном соке (4...5 мг%) (Björck L., 1982).
Активирование лактопероксидазо-антибактериальной системы молока имеет немаловажное значение для сыроделия. Были проведены исследования, в которых молоко инокулировали Ps. fluorescens и выдерживали при 4 °С в течение 3 сут. Бактериальная обсеменность контрольного (необработанного) молока увеличилась через 2 сут в 10 раз, через 3 сут – в 100 раз. В молоке с активированной лактопероксидазой количество микроорганизмов сократилось в 100 раз. Имеются данные, что при хранении сырого молока при (3 ± 1)°С в течение 8 сут, в которое через 1, 3 и 5 дней хранения было добавлено по 1,5 мг% SCH- и Н2О2 не происходило существенных изменений обсеменения молока психрофильными бактериями и его вкуса по сравнению с контролем. При этом выход сыра увеличивался на 2 % по сравнению с выходом сыра при использовании необработанного молока (Zall R. R., Chen J. Н., Dzuree D. J., 1983). Кроме чисто антибактериального эффекта, лактопероксидазо-антибактериальная система молока инактивирует некоторые ферменты, участвующие в гликолизе молока, и тем самым способствует снижению количества молочной кислоты, образующейся при гликолизе молока в процессе его хранения.
При добавлении к цельному сырому молоку по от 0,03 до 0,3 ммоля натрия тиоцианата и натрия перкарбонатаи хранении такого молока при 5 °С в течение от 6 до 72 ч отмечалось постепенное заметное ингибирование липопротеинлипазы (в среднем на 30 %), однако при этом явного снижения липолиза молока, по сравнению с контролем, не наблюдалось, что можно объяснить защитной ролью ОШЖ (Ahrné L., Björck L., 1985).
Другим путем повышения активности лактопероксидазо-антибактериальной системы молока при его хранении является использование различных оксидаз в иммобилизованном виде, например глюкозооксидазы (с добавлением или без добавления незначительных количеств глюкозы – 0,1...0,5%), аминоксидазы (с добавлением небольших количеств глицина – 0,5%), ксантиноксидазы (с добавлением гипоксантина) и других, которые, воздействуя на соответствующие субстраты, повышают в молоке содержание перекиси водорода (Fox P. F, 1980). При этом количественный состав тех или иных вносимых в молоко компонентов зависит от условий, которые имеются или которые необходимо создать для улучшения бактериального исходного состояния молока, а также и от исходного качества молока.
В районах с жарким климатом указанные пути стабилизации качества сырого молока могут быть использованы для уменьшения случаев порчи молока при доставке его из отдаленных мест, а также увеличения ресурсов молока, пригодного для производства высококачественных молочных продуктов.
Показано отсутствие влияния наличия в молоке соматических клеток в количестве 200...1500 тыс/мл на действие лактопероксидазо-антибактериальной системы молока (Гудков А. В., 1985).
После 15 лет экспериментов в полевых условиях в развитых и развивающихся странах Инструкция по использованию альтернативного метода консервирования молока, основанная на активировании натурального антибактериального комплекса в молоке (ММФ, 1988, № 234), одобрена Комитетом экспертов по пищевым добавкам ФАО/ВОЗ в 1989 г. и Комиссией Кодекса Алиментариус в 1991 г. (Бюл. ММФ, 2001, № 365; Ламберт Д. С, 2003).
При охлаждении до достаточно низких положительных температур и хранении такого охлажденного молока происходит некоторое изменение агрегатного состояния триглицеридов молочного жира, в результате чего нарушается целостность структуры ОШЖ, которая становится хрупкой и несколько проницаемой для жидкого свободного жира. Перемешивание увеличивает эту степень дестабилизации, а свободный жир подвергается действию как нативных липаз, так и заметно — липазами психрофильных бактерий при условии их содержания ≥ 106... 107 КОЕ/мл.
Широкое применение машинного доения и хранения молока, охлажденного до достаточно низких температур, в течение длительного периода до его тепловой обработки ставит перед молочной промышленностью проблему липолиза, следствием которого являются развитие в молоке специфического липолизного (липолизированного) вкуса и ухудшение ряда технологических свойств молока. Чем дольше молоко хранится, тем больше возможность развития в нем микробного липолиза, особенно в результате жизнедеятельности психрофильной микрофлоры, обладающей высокой липолитической активностью (гл. 1). При длительном хранении молока в условиях достаточно низких температур повышается суммарная липолитическая активность молока, результатом чего является увеличение содержания в молоке продуктов гидролиза молочного жира (рис. 34).
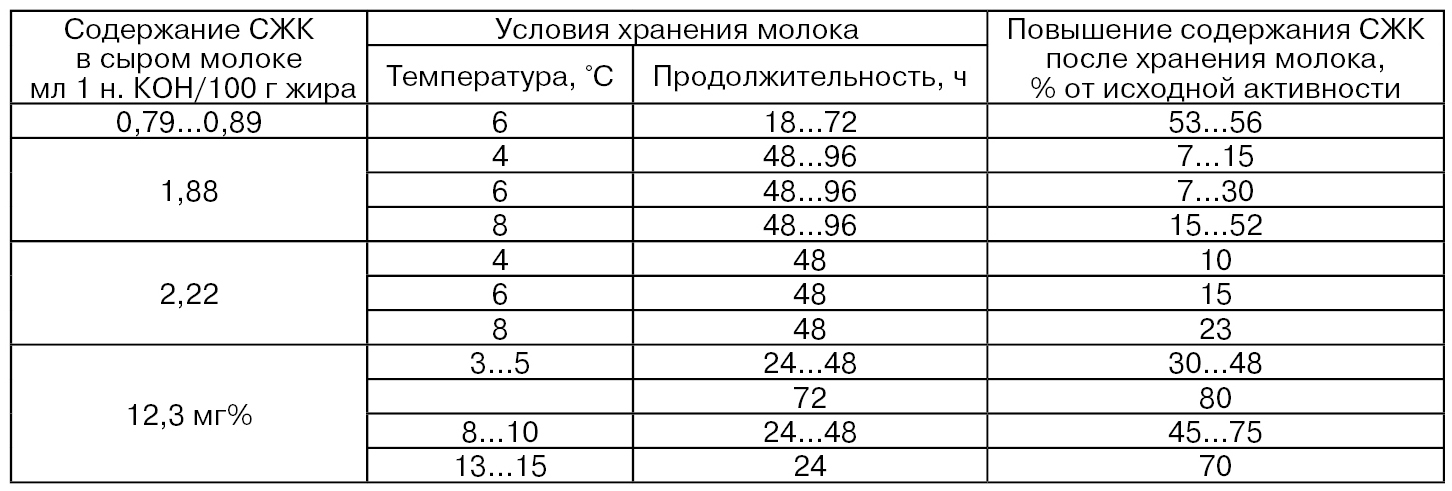
Данные табл. 40 показывают, что при различных условиях хранения сырого молока повышение содержания СЖК составляло от 7 до 80 % в зависимости от исходной величины (Шидловская В. П., Патритий А. П., Насонова Л. М.; Downey W. К., 1980).
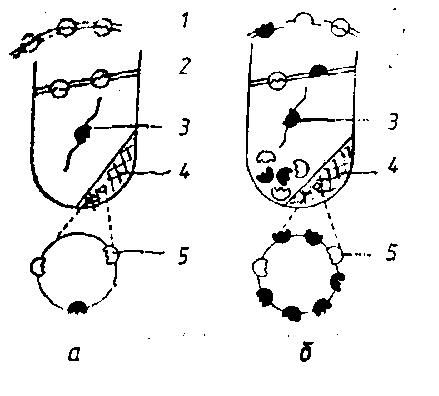
Рис. 34. Изменение локализации нативной липопротинлипазы (ЛПЛ) при хранении сырого молока при 4 °С в течение 24 ч (Downey W. К., 1980):
а – до хранения; б – после хранения;
1 – оболочка шариков жира; 2 – сливочный слой (центрифугирование цельного молока при 10 000 g в течение 20 мин при 4°С); 3 – активная форма ЛПЛ; 4 – осадок казеина; 5 – неактивная форма ЛПЛ
Результаты хранения сборного сырого охлажденного молока (ежемесячно, в течение года от около 2000 молочных ферм) с различным уровнем бактериальной обсемененности – от ≤ 300 тыс. до > 3 · 106 КОЕ/мл в течение трех суток показали, что содержание СЖК составило в среднем 0,52; 0,68 и 0,75 мэкв./л соответственно без заметных различий в бактериальной обсемененности в первые и вторые сутки хранения и заметных различий – в третьи сутки хранения, а именно – от 0,65 до 0,85 мэкв./л (Suhren G., Heeschen W, 1991). Таким образом, нельзя исключать влияния микробного липолиза при хранении сырого охлажденного молока через 2...3 сут на его качество.
Таблица 40
Показано, что при хранении сырого молока от отдельных коров при 4°С в течение 3 сут активность липопротеинлипазы постепенно увеличивалась с 40,4 ед./мл – в контроле до 57,4 ед./мл – в концу хранения. При этом в основном за счет освобожденной липопротеинлипазы из казеиновой фракции молока – с 29,9 в контроле до 44,4 ед./мл (Hohe К. А., Dimick Р. S., Kilara А., 1985). При хранении сборного молока, по сравнению с молоком от отдельных коров, повышение активности липопротеинлипазы выражено в большей степени также за счет большего освобождения фермента из казеиновой фракции молока, а именно 43,1 и 32,3 % соответственно (Anderson М., 1982).
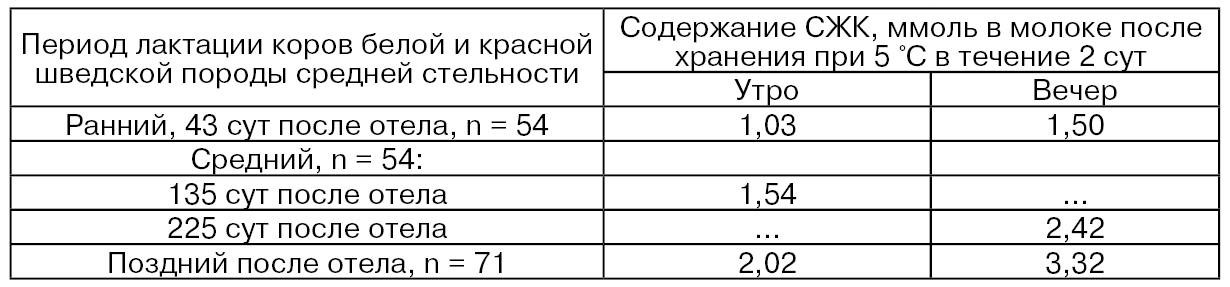
Величины активности липопротеинлипазы в хранении молока зависят от стадии лактации и времени удоя, о чем свидетельствуют приведенные в табл. 41 данные (n – количество коров) (Ahrné L., Björck L., 1985).
Таблица 41
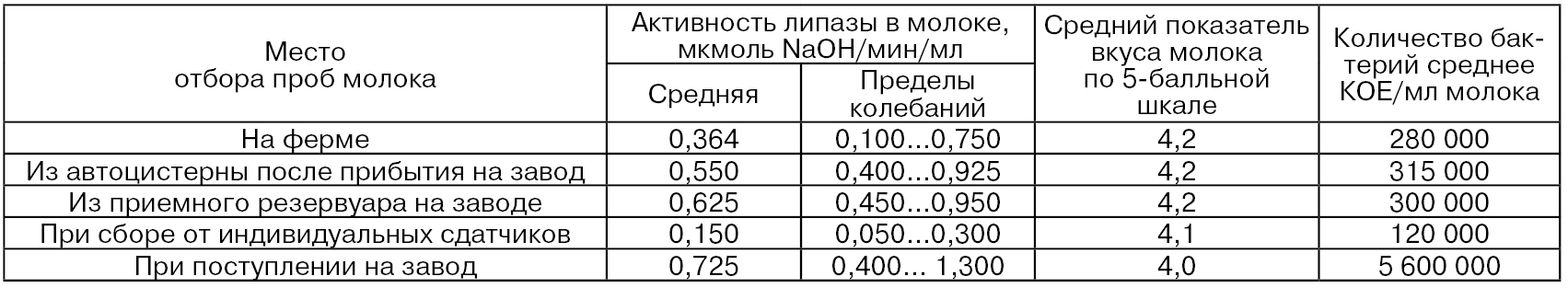
Вследствие механического воздействия на молоко при перекачивании и взбалтывании во время транспортирования активность липазы также повышается. Как видно из табл. 42, при транспортировании сборного молока с фермы на завод активность липазы повышается на 51 %, а при сборе от индивидуальных сдатчиков – на 400 %, так как сбор молока от индивидуальных сдатчиков происходит через день, а зимой даже через 2 или 3 дня.
Таким образом, по различным причинам при ферментативном гидролизе молочного жира в молоке накапливаются моно- и диглицериды, СЖК, В дальнейшем происходит ферментативное окисление СЖК. При окислении предельных жирных кислот образуются различные кетоны (например, метиламилкетон). При окислении непредельных жирных кислот промежуточными продуктами являются окси- и кетокислоты, конечными продуктами могут быть ди- и монокарбоновые (например, пеларгоновая) кислоты, углеводороды. В результате окисления жирных кислот с сопряженными и несопряженными двойными связями (линолевая, линоленовая и арахидоновая) с участием липоксигеназы образуются неустойчивые и высокоактивные гидроперекиси и перекиси, а в дальнейшем – альдегиды, кетоны, кислоты, оксисоединения и другие вещества, обладающие неприятным запахом и вкусом. Образование различных продуктов гидролиза молочного жира может привести к снижению поверхностного натяжения молока и развитию прогорклого, липолизированного привкуса, что снижает качество сливок, масла и других молочных продуктов, выработанных из такого молока (Шидловская В. П., Аристова В. П., 1981; Anderson М., 1987; Nicholson J. W. G., 1993).
Таблица 42
При исследовании молока, в которое вносили чистые культуры психрофильных бактерий и выдерживали при 4°С, установлено, что 17 видов из 25 вызывали появление прогорклого вкуса, при этом их активность была различной по дням возникновения этого порока молока, а именно от 4 до 12 сут. Органолептические изменения молока под действием липолитических ферментов наблюдается, когда число активных растущих психрофилов достигает от 106 до 108 КОЕ/мл.
Изменение химического состава молочного жира в результате его ферментативного гидролиза приводит к нарушению параметров технологических режимов производства молока и молочных продуктов. Например, вследствие снижения поверхностного натяжения молока увеличивается продолжительность сбивания сливок; при частичном изменении дисперсности жировой фазы молока уменьшается степень обезжиривания молока при сепарировании; в результате угнетающего действия СЖК на молочнокислые бактерии увеличивается продолжительность сквашивания молока и сливок, а также продолжительность сычужного свертывания; уменьшается продолжительность хранения молока и молочных продуктов.
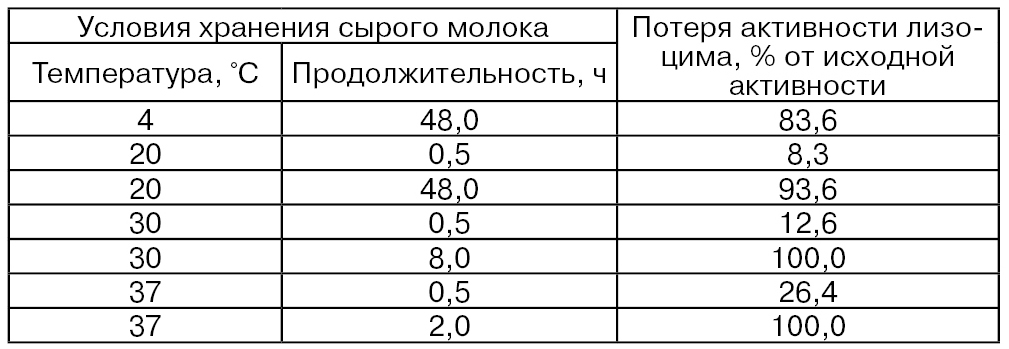
При хранении молока происходит потеря активности антибактериального фермента лизоцима (табл. 43) (Shahani К. М., Chandan R., Kelly Р.). Эти потери тем больше, чем выше температура и продолжительнее хранение молока.
В результате проявления активности протеолитических ферментов как нативного, так и микробного происхождений при хранении сырого молока в условиях достаточно низких температур происходит протеолиз (по Reimerdes E. Н., 1983).
При хранении сырого молока в условиях достаточно низких положительных температур гидрофобные взаимодействия в мицеллах казеина ослабевают, что способствует выходу из них β-казеина, фосфата кальция и, наряду с нативными липазами, щелочной протеиназы. Так, при хранении сырого молока в течение 48 ч при 4 °С содержание растворимого казеина (основную часть которого составляет β-казеин – важнейший стабилизирующий компонент мицеллы казеина) постепенно увеличивается почти на 60 %, в то время как при 10 и 15 °С его содержание снижается на почти 13 и 23 % соответственно по отношению к контролю – 4,4 мг/мл. Диссоциированный в водную фазу молока β-казеин подвергается гидролизу нативными и микробными протеиназами с образованием γ-казеинов и фосфопептидов. Например, содержание γ-казеина при хранении молока в течение 24 ч при 4 и 11 °С может повыситься почти на 25 и 15 % по сравнению с исходным содержанием (рис. 35) (Reimerdes Е. Н., Herlitz Е., 1979). При этом следует отметить, что γ-казеин не свертывается сычужным ферментом и при производстве творога и сыра уходит с сывороткой, что снижает выход этих продуктов
Таблица 43
В то же время, результаты других исследований показывают, что гидролиз β-казеина с образованием γ-казеинов при повышенных температурах хранения молока идет в несколько раз быстрее, чем при 4°С. Кроме того, не обнаружено потерь в активности плазмина мицелл казеина, полученных из молока, хранившегося при 4 °С в течение 16 ч, по сравнению с молоком при 20°С (Grufferty М. В., 1986).
При хранении молока возможен переход неактивной формы щелочной протеиназы I в активную форму, что обусловлено наличием в молоке активаторов этого фермента (Каnу J.J., Frank J. F., 1988).
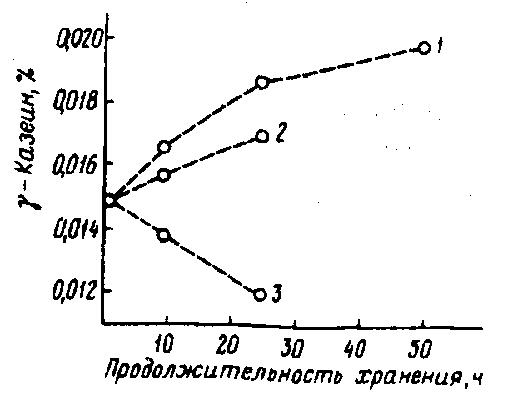
Рис. 35. Изменение содержания γ-казеина в зависимости от продолжительности и температуры хранения сырого молока:
1 – 4°С; 2 – 11°C; 3 – 26°С
Наибольшее участие в протеолизе молока принимают протеолитические ферменты психрофильной микрофлоры. Следует отметить, что после 2...3 сут хранения молока при 4...8°С различие между содержанием общего количества бактерий и психрофильных микроорганизмов незначительное. В этот период число психрофильных бактерий достигает 106 КОЕ/мл, и они становятся особенно активными продуцентами не только липолитических, но и протеолитических ферментов и могут стать существенной причиной порчи молока.
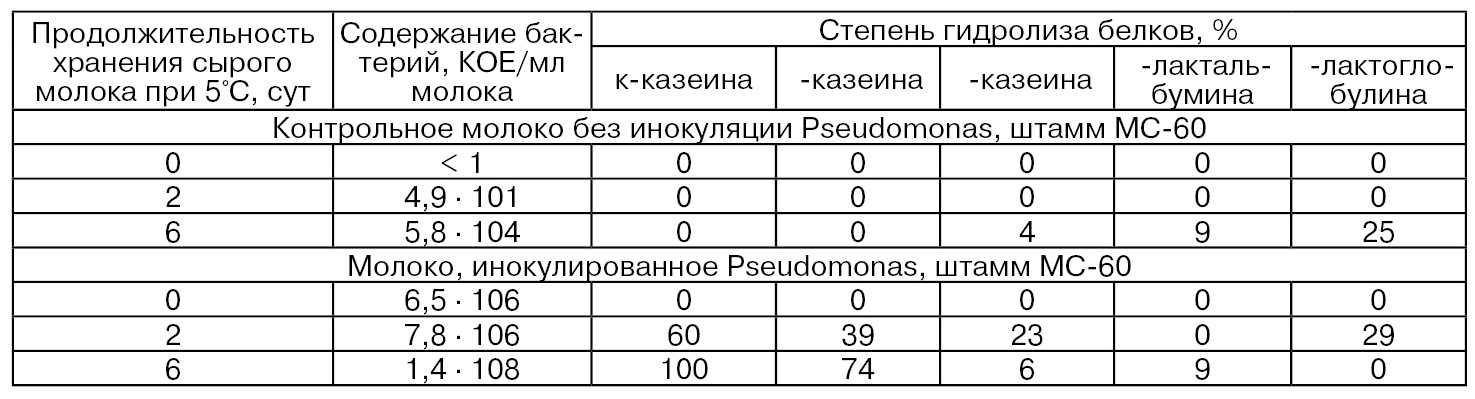
В табл. 44 приведены данные о влиянии протеаз психрофильных бактерий рода Pseudomonas на гидролиз различных фракций казеина и сывороточных белков при хранении сырого обезжиренного молока (Adams D. М., Barach J. Т., Speck М. L.). При этом исходное сырое молоко получали в асептических условиях, затем быстро охлаждали до 4°С, хранили в течение суток и сепарировали. Полученное обезжиренное молоко засевали Pseudomonas (штамм МС-60) в количестве 106 КОЕ/мл (культуру предварительно выделяли из сырого молока, хранили при 2°С в стерильном обезжиренном молоке и затем 0,1 мл культуры засевали в восстановленное стерильное обезжиренное сухое молоко и инкубировали 24 ч при комнатной температуре).
Таблица 44
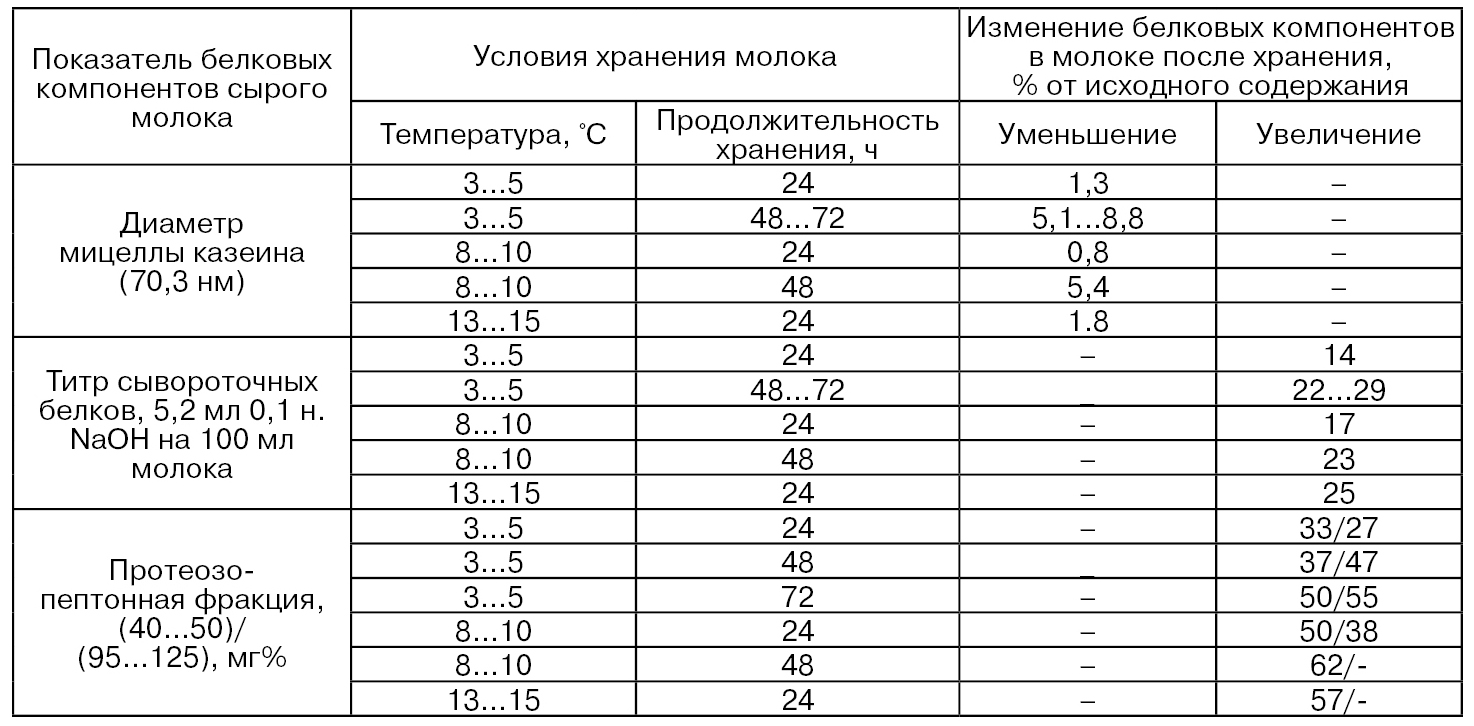
Вследствие активности протеолитических ферментов изменяются диаметр мицелл казеина, титр сывороточных белков и содержание протеозо-пептонной фракции (табл. 45) (Шидловская В. П., Патратий А. П.).
Таблица 45
В результате гидролиза основных фракций белков молока снижается его питательная ценность. В молоке происходит накопление пептидов, в том числе обладающих горьким вкусом, и различных небелковых азотистых соединений, например аминокислот, которые в дальнейшем при тепловой обработке могут участвовать в реакции Майяра (побурение молока).
Установлено, что уменьшение содержания основных белковых фракций молока при ферментативном гидролизе вызывает увеличение продолжительности сычужного свертывания молока, снижение прочности сгустка и тем самым обусловливает ухудшение качества сыра. Сгущенное молоко, выработанное из долго хранившегося молока, имеет вязкую консистенцию и невыраженные вкус и запах. Снижение содержания к-казеина может стать одной из причин коагуляции белков при высокотемпературной обработке молока или вскоре после нее.
Важным фактором в развитии липолитической и протеолитической активностей психрофильной микрофлоры при хранении сырого молока является состояние молочного оборудования. Так, если при тщательной обработке молочного оборудования количество психрофильной микрофлоры через 3 ч после доения составило от 7,0 до 2,0 · 102 и после хранения при 3...5 °С в течение 72 ч – в среднем 6,3 · 103, то при недостаточной обработке этого оборудования – от 6,1 · 103 до 1,3 · 104 и от 1,5 · 106 до 2,4 · 107 соответственно, т. е. на уровне, при котором происходит наибольшее образование экзоклеточных липолитических и протеолитических ферментов (Моисеева Е. Л., 1988).
Таким образом, в зависимости от условий получения и первичной обработки сырое молоко при доставке на молочное предприятие будет иметь различную по виду и величине ферментативную активность. В целях предотвращения или снижения возможных ферментативных изменений сырого молока при его получении и первичной обработке необходимо обеспечивать адекватное кормление коров; уделять особое внимание вопросам санитарно-гигиенического состояния ферм; избегать большого количества коров в стаде в конце лактации; снижать уровень мастита; строго соблюдать правила монтажа и эксплуатации доильных установок; для снижения степени агрегирования шариков жира молока при прохождении его по молокопроводу у стенок следует поддерживать температуру не менее 35 °С, для этого через молокопровод перед доением коров необходимо пропускать горячую воду (50...60°С); максимально снижать возможность подсоса воздуха через доильный аппарат, соединения молокопровода, вакуумный фильтр и др.; охлаждать молоко с момента его получения до 5...6 °С в течение не более 2 ч; скорость движения потока молока в молокопроводах следует поддерживать на уровне 1...2 м/с; не допускать интенсивного перемешивания парного молока в течение длительного времени; не допускать смешивания парного молока с охлажденным молоком, а также периодического нагревания и охлаждения молока; максимально снижать продолжительность перемешивания молока в резервуарах и не допускать многократного перекачивания молока насосами (не более 3 раз); дифференцировать температуру охлаждения молока в зависимости от механических воздействий, которым молоко было подвержено до охлаждения (чем больше молоко было подвержено таким воздействиям, тем температура его охлаждения должна быть ниже); обеспечивать соответствующую мойку и дезинфекцию доильного оборудования, емкостей для хранения молока и автоцистерн; общая продолжительность хранения молока на фермах и его транспортирования не должна превышать 48 ч при 4...6°С и 12...24 ч при 8...10°С в зависимости от исходного микробного обсеменения молока. Результаты исследований показывают, что при хранении сырого молока в атмосфере азота при 4 °С (расход азота 100 мл/мин) значительно замедляется развитие психрофильных бактерий. С этой же целью рекомендуется осуществлять мягкую тепловую обработку – термизацию сырого молока на ферме сразу после доения или непосредственно после приемки молока на заводе при температуре 60...65 °С с выдержкой 10...25 с, после чего его охлаждают до температуры ниже 8 °С и направляют на хранение. Показано, что термизация способствует гибели большинства психрофильных бактерий (может подавить их развитие на 85%) еще до того, как они стали бы развиваться и быть источником термостойких экзолиполи- и протеолитических ферментов (Храмцов А. Г., Емельянов С. А. и др., 2006). В тех же случаях, когда в молоке содержание психрофильной микрофлоры достигло миллиона или более КОЕ/мл, с целью инактивации уже накопленных при жизнедеятельности этой микрофлоры протеолитических и других ферментов следует увеличить продолжительность первичной тепловой обработки молока от нескольких минут до 1 ч, при этом температуру обработки можно снизить до 50...55 °С (п. 3.1), что позволит значительно снизить возможность развития ферментативных реакций окисления, липолиза и протеолиза в пастеризованном молоке.
Этим вопросам большое внимание уделяется в периодических изданиях ММФ.
[1] Björck L., 1978; Янковский Д.С., Попова Н.Г, 1987; Gönс S., Akbulut N. et al., 1990; Бюл. ММФ, 1991, №264; Димитров Т. и др., 1992; Zapico Р., Gaya et al., 1992; Бюл. ММФ, 1993, №284; Barrett N.E. et al., 1999; Zarung S.A., Gnan S.O., 1999.


