




Целый ряд зоотехнических факторов, включающих породу коров, период лактации и молочную продуктивность, кормление, условия содержания, индивидуальные особенности, физиологическое состояние и заболевания коров, а также время года, играют существенную роль в ферментативной активности молока. Приспособление животных к различным условиям содержания и кормления сопровождается изменением их обмена веществ, обусловленным наряду с другими факторами изменением ферментативных систем организма, в том числе молочной железы. Для некоторых ферментов молока имеют значение и генетические факторы. В связи с указанным, изучение влияния различных зоотехнических факторов на изменение содержания и активности ферментов молока представляет определенный интерес.
Порода коров. Имеются определенные различия в содержании и активности отдельных ферментов молока в зависимости от породы коров. Из табл. 18 видно, что, например, активность ксантиноксидазы и щелочной фосфатазы в молоке отдельных пород коров может различаться до 2 раз. Более выражена зависимость активности липазы молока от породы коров. Различие в этих активностях составляет до 3,5 раз.
Различие в активностях ферментов в молоке в зависимости от породы коров обусловливается их молочной продуктивностью, условиями содержания и кормления.
Период лактации и молочная продуктивность коров. Продолжительность лактации у коров составляет в среднем 300 дней. Ее подразделяют на три периода: молозивный период (7...10 дней после отела), период секреции нормального молока (285...277 дней) и период секреции стародойного молока (5...10 дней перед сухостойным периодом). По химическому составу молозиво и стародойное молоко значительно отличаются от нормального молока. Так, в них больше белка, жира, меньше лактозы, больше биологически активных веществ, в том числе и ферментов. В соответствии с ГОСТ Р 52054 «Молоко натуральное коровье-сырье», молозиво и стародойное молоко не подлежат приемке на пищевые цели.
Таблица 18
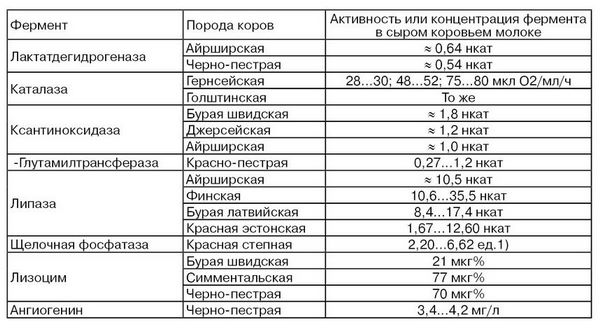
Примечания. Активность плазмина ниже в молоке коров джерсейской породы и выше в 2 раза в молоке фризской, чем айрширской пород. Концентрация пероксид-дисмутазы на 25% больше в молоке джерсейской породы, чем в молоке голштинской.
1) 1 ед. соответствует количеству фермента, вызывающего образование 1 мкг фенолфталеина/мл молочной сыворотки в течение 1 ч при 37 °С.
С началом лактации в ферментативной системе молочной железы происходят соответствующие адаптивные изменения. Например, в лактирующей молочной железе, по сравнению с нелактирующей, активность дегидрогеназ повышается почти в 2 раза, щелочной фосфатазы – в 6 раз, аргиназы – в 1,5 раза. Отмечается и некоторое повышение активности ксантиноксидазы, кислой фосфатазы и протеиназ (И. А. Шумкова). Поэтому важно определение зависимости между периодом лактации, связанной с ним молочной продуктивностью животных и активностью в молоке ферментов. Величина этой зависимости свидетельствует о степени участия тех или иных ферментов в синтетических процессах молочной железы и, следовательно, может служить тестом молокообразования. Активность целого ряда ферментов в молозиве и нормальном молоке резко различается (табл. 18).
Данные, приведенные в табл. 19, показывают, что примесь молозива в нормальном коровьем молоке может привести к повышению в нем активности таких основных ферментов, как лактопероксидазы, арил- и холинэстераз, кислой фосфатазы, рибонуклеазы, α-амилазы, протеаз и др.
Таблица 19
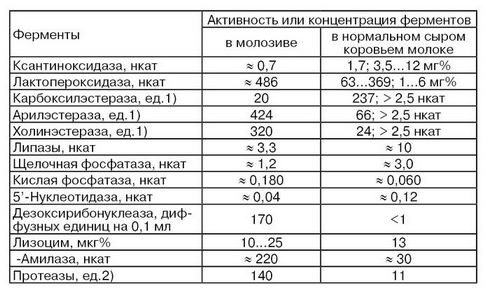
1) 1 ед. соответствует количеству фермента, которое катализирует выделение 1 мл СО2 в 1 мл молока в течение 30 мин при принятых условиях реакции.
2) 1 ед. соответствует количеству фермента, вызывающего образование 1 мкг тирозина в 1 мл молока.
В ходе лактации в связи с изменением уровня молочной продуктивности активность многих ферментов в молоке изменяется. Так, при повышении удоев коров с 10 до 18 л активность лактатдегидрогеназы в молоке снижалась с 0,09 до 0,06 Е (с ≈ 1,5 до ≈ 1,0 нкат). В течение лактации активность этого фермента снижается, но несколько повышается к концу лактации. С увеличением числа лактации коров (с 1 до 6) активность лактатдегидрогеназы в молоке также несколько повышается.
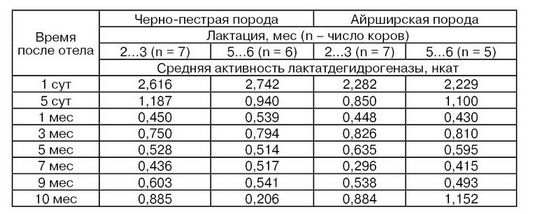
Динамика активности лактатдегидрогеназы в течение лактации тесно связана с изменением ее субстратов в молоке (Грачев И. И., Попов С. М., Скопичев В., Т. Sonneck R., 1979). В табл. 20 показано изменение активности этого фермента в течение лактации, в молозиве и молоке коров черно-пестрой и айрширской пород (Курдуманова О. И., 1987). В первые сутки после отела активность фермента максимальна, и к концу первого месяца лактации она снижается в 5...6 раз. На третьем месяце лактации активность этого фермента в молоке заметно увеличивается, что связано с началом стельности, затем организм коровы адаптируется и активность фермента снижается, а к концу лактации, при подготовке к новому отелу, активность этого фермента значительно повышается. Следует отметить, что изоферментный спектр лактатдегидрогеназы также подвержен изменениям в ходе лактации. Так, если сразу после отела в молозиве коров 2...3 лактации выявлено 5 изоферментов, то у коров 5...6 лактации обнаружено только 3 изофермента, формы 4 и 5 отсутствовали. К 5 сут в молоке этих коров оставалось 2 изофермента, а в молоке коров 2...3 лактации – 3.
Таблица 20
Что касается ксантиноксидазы, то в течение месяца после отела коров активность этого фермента в молоке сохраняется на довольно постоянном уровне (с колебаниями в 5...6 %). Однако в целом за время лактации активность ксантиноксидазы увеличивается. По данным И. А. Шумковой, за 8 мес лактации коров холмогорской породы активность ксантиноксидазы в молоке повышается в 2...4 раза. При этом более высокая активность фермента отмечена в молоке высокопродуктивных коров (от 5000 до 8000 л), чем в молоке низкопродуктивных коров (до 3000 л). На протяжении периода лактации отмечена корреляция между активностью ксантиноксидазы и суточным удоем молока (коэффициент корреляции 0,41...0,98).
Довольно заметно повышается в молоке к концу лактации активность каталазы. Это можно, видимо, объяснить заметным увеличением числа соматических клеток в этот период, которые характеризуются высокой каталазной активностью. В то же время активность лактопероксидазы в молоке в течение лактации снижается.
Из трансфераз следует отметить γ-глутаминтрансферазу, содержание которой значительно зависит от стадии лактации, а именно наиболее высокое в молозиве с постепенным снижением в течение двух недель лактации (Kitchen В. J., 1985).
В связи с особой ролью, которую играет липолиз в снижении качественных показателей молока, вопросы, связанные с влиянием на степень его выраженности некоторых зоотехнических факторов, очень важны.
Так, снижение линейной корреляции между активностью липопротеинлипазы и степенью липолиза установлено в сливках, полученных из сырого молока (при центрифугировании цельного молока при 500 g в течение 20 мин при 4°С), в зависимости от периода лактации. Коэффициент корреляции составил 0,91; 0,82 и 0,71 соответственно в течение 1,5 и 4 мес лактации (Sundhein G., Bengtsson-Olivecrona G., 1985; Chavarri F., Santisteban A. et al., 1998).
Показано, что липаза молока и молозива различаются между собой по молекулярной структуре. После исчезновения липолитической активности в молозиве, четко обнаруживается активность липазы молока. Следовательно, возможно, молозивная липаза является проферментом молочной липазы. Молозивная липаза образуется в течение первых двух дней после отела или, может быть, перед отелом, и она не играет какую-либо важную роль в развитии липолиза. Эта липаза не связана с казеином, не активируется сывороткой крови, но активируется при перемешивании (Driessen F. М.).
Отмечено некоторое различие в липолитической активности молока различных доек. Так, на ранней стадии лактации коров в молоке вечерней дойки эта активность выше, чем в молоке утренней дойки. Кроме того, чем выше надой молока, тем ниже его липолитическая активность (Ahrne L., Björck L., 1985).
Установлено, что молоко последней стадии лактации особенно чувствительно к спонтанному липолизу. Поэтому если в одно и то же время большое количество коров на ферме находится на последней стадии лактации, то это может представить большую опасность появления «липолизированных» пороков вкуса и запаха молока. Так, из рис. 22 видно, что на последней стадии лактации коров, а также при снижении количества надоенного молока и увеличении общего числа лактации в молоке значительно повышается содержание СЖК.
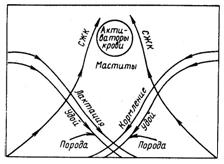
Рис. 22. Факторы, влияющие на развитие спонтанного липолиза в сыром молоке (Downey W. K., 1980)
Данные, приведенные на рис. 23 по изменению активности липопротеинлипазы в течение лактации 12-ти коров и соответствующих величин кислотного числа (ADV), показывают, что активность фермента, начиная с первого месяца лактации, постепенно увеличивается, достигая максимума к пяти месяцам лактации, а количество образующихся СЖК – к семи месяцам.
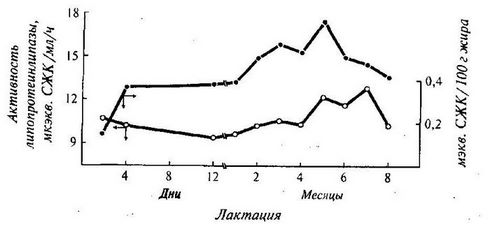
Рис. 23. Изменение активности липопротеинлипазы в молоке в течение лактации (по Chilliard Y., 1982)
В молозиве первых суток отмечается высокий уровень активности фосфатаз, что, вероятно, объясняется накоплением их в молочной железе перед отелом и увеличением мембранной проницаемости клеток. Увеличение активности кислой фосфатазы в молоке на третьем месяце лактации, а щелочной – на втором месяце можно объяснить высокими надоями молока в эти периоды. К концу лактации, перед новым отелом под влиянием гормонов изменяется клеточный состав молочной железы с одновременным увеличением активности щелочной и кислой фосфатаз, наряду с лактатдегидрогеназой (Валова Л. В., Курдуманова О. И., 1988).
По данным многих исследований, активность щелочной фосфатазы в течение лактации находится в пределах 1800...4700 мкг фенола/мл или 500...1000 мкг n-нитрофенола/мл/30мин (Harding F., 1991). Биологическое варьирование щелочной фосфатазы в сыром сборном молоке составляет 25 %, а между отдельными коровами – достигает одного порядка (Schlimme Е., Thiemann А., 1992).
Отмечают, что в течение лактации активность щелочной фосфатазы повышается значительнее, чем кислой (соответственно на 108...829 и 57...367 %), Повышение активности фосфатаз в молоке высокопродуктивных коров, в особенности кислой фосфатазы, происходит медленнее, что можно объяснить более плавным падением у них суточных удоев. Высокая активность щелочной фосфатазы, как и липазы, в молоке коров отмечается перед сухостойным периодом. Установлено, что всякое изменение удоя молока связано с обратным изменением активности фосфатаз. Например, коэффициент корреляции между активностью в молоке коров кислой фосфатазы и суточным удоем в среднем за 8 мес лактации составил – 0,60, а в конце лактации – 0,90 и 1,0 (И. А. Шумкова, К. А. Порядкова, С. Н. Любчик). По данным Linden G., Ged J. (1978), повышение активности щелочной фосфатазы в молоке коров в течение лактации происходит неравномерно. Если в течение первых 20 недель лактации активность фермента в молоке постепенно повышалась с 10 до 40...50 ед. (1 ед. соответствует количеству фермента, вызывающего образование n-нитрофенола, мкмоль/мин/л), то в остальные 20 недель активность его возросла до 200...280 ед. Одновременно отмечается постоянное снижение активности фермента в молоке с увеличением числа лактации коров. Так, если активность щелочной фосфатазы в молоке коров первой лактации находилась в пределах 35...107 ед., третьей лактации – 14...55 ед., то в молоке пятой лактации – только 16...19 ед.
Адаптивные изменения активности ферментов молока в связи с уровнем молочной продуктивности коров подтверждают результаты работ Л. М. Буруянэ, А. С. Георгиу (1978), показавших, что число изоформ лактопероксидазы в молоке находится в обратной зависимости, а щелочной фосфатазы — в прямой зависимости от молочной продуктивности коров. В отличие от этого число изоформ кислой фосфатазы высоко- и среднепродуктивных коров сходно.
Высокая активность дезоксирибонуклеаз определена в молозиве различных сельскохозяйственных животных и, в ряде случаев, в несколько раз выше, чем в крови. Дезоксирибонуклеаза коровьего молозива по некоторым свойствам близка к этому же ферменту селезенки и отличается от таковой поджелудочной железы коров. Оптимумы рН дезоксирибонуклеаз коровьего молозива составляет 5,0...5,5 (Адигамов Л. Ф., 1986).
Приведенные в табл. 21 данные по содержанию рибонуклеазы и ангиогенина показывают, что в молозиве первых суток отмечается наиболее высокий их уровень. В дальнейшем их содержание постепенно снижается с приближением к средним для нормального молока значениям концентраций. При этом превышение уровня рибонуклеазы и ангиогенина в молозиве, по сравнению с нормальной секрецией, выражено менее резко, чем для общего белка. Это может быть связано с тем, что эти ферменты попадают в молоко из крови, в то время как значительное повышение уровня общего белка в молозиве может быть связано с белками из молочной железы, в которой в начальный период лактации происходят адаптивные изменения. В стародойном молоке, на фоне некоторого повышения общего белка, отмечается тенденция к увеличению количества рибонуклеазы и ангиогенина. Показано, что коровы (на примере черно-пестрой породы) после первого отела продуцируют молоко с содержанием ангиогенина в среднем на 20% меньше, чем после нескольких отелов, а изменение содержания в молоке ангиогенина в течение лактации, например, коров голштино-фризской, черно-пестрой, холмогорской и айрширской пород подобно (Шалыгина А. М., Ионова И. И. и др., 1997; Тихомирова Н. А., Комолова Г. С, Ионова И. И., 2004).
Таблица 21
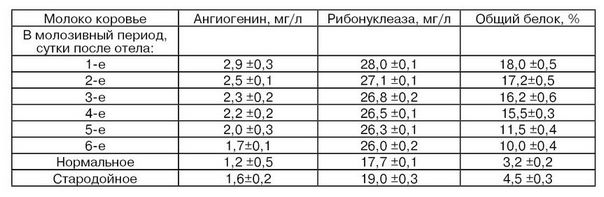
Для молозива характерно следующее распределение активности изоформ рибонуклеазы:
|
Форма рибонуклеазы |
Активность, ед[1]/мл |
|
А |
14000(67) |
|
Б |
6000 (20) |
|
II-1 |
780(4) |
По активности в молозиве форма II-1 в 10...15 раз превышает активность ее в молоке, что свидетельствует о ее значительной роли в иммунитете организма животного в постнатальный период (Meyer D. Н. et al, 1987).
Результаты исследований И. Ф. Лысенко (1980) показывают, что активность амилаз в молоке в среднем по месяцам лактации колеблется от 16,8 до 35,2 амилазных единиц. Отмечена невысокая зависимость между среднесуточным удоем коров и активностью амилаз в молоке (коэффициент корреляции +0,53).
В зависимости от стадии лактации, содержание лизоцима в молоке находится в пределах 10...32 мкг%. Наиболее выраженная активность лизоцимов типов М, Т и Мз в молоке отмечается в период 2...6 мес лактации, т. е. в период наиболее высоких удоев коров (рис. 24). Отмечают, что в молоке утреннего удоя концентрация лизоцима выше, чем в молоке вечернего удоя (Shahani К. М., Chandan R., Kelly P.).
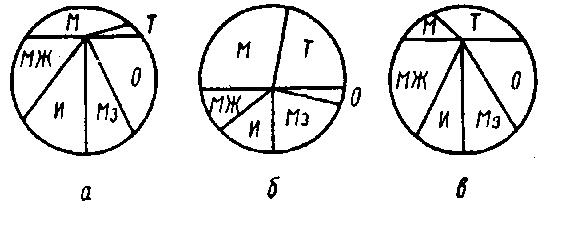
Рис. 24. Относительная активность лизоцима в молоке, полученном от здоровых коров в течение лактации (Мутовин В. И.):
а — 4...10 сут; б — 2...6 мес; в — от 7 мес до конца лактации; М, Т, МЖ, И, Мз, О — типы лизоцима
В табл. 22 приведены данные по активности лизоцима типа М к некоторым болезнетворным бактериям, в зависимости от периода лактации коров (Р. А. Бабаев), которые показывают, что в первые 3 мес после отела активность лизоцима типа М наиболее высокая, а затем снижается. При этом, она была более выражена к грамположительным и менее к грамотрицательным бактериям.
Таблица 22
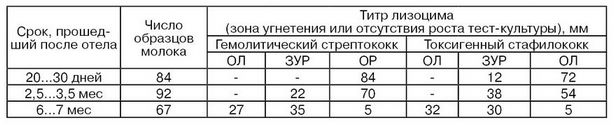
Обозначения: ОЛ – отсутствие лизоцима в молоке; ЗУР – зона угнетения роста в пределах 20...45 мм; ОР – отсутствие роста в пределах 20...45 мм.
Отмечено, что количество лизоцима в молозиве в 30 раз больше, чем в сыворотке крови. Фермент или избирательно поглощается молочной железой из крови, или синтезируется в ее клетках. Возможно, его накопление в молозиве осуществляется по двум указанным путям (по Горбатовой К. К., 2004).
Исследования по важнейшему из гликозидаз молока ферменту N-ацетил-β-глюкозаминидазы показали, что секреция в сухостойный период в 10 раз превышала ее уровень в обычном молоке. Стадии стельности мало влияли на активность этого фермента. Обнаружена значительная зависимость между стадией лактации и активностью этого фермента. Так, она была выше в начале, снижалась в середине и увеличивалась в конце лактации и в период сухостойной секреции. Увеличение активность фермента в молоке в первую стельность коров в конце лактации оказалась менее значительной, чем во вторую и третью лактацию (Miller R. Н., Рааре М. J., 1988).
Результаты анализов около 400 проб сырого молока коров голштинской и джерсейской пород, полученного в течение 10 мес лактации, показали, что активность плазмина увеличивалась в течение лактации, но не зависела от породы, pH и состава молока (наибольшее влияние на нее оказывало количество лактации). Активность плазминогена зависела только от стадии лактации и сезона года. Наиболее высокая его активность отмечена в период с 4-го месяца и до конца лактации, а также осенью и зимой (Bastian Е. D., Brown R. J., Ernstrom С. А., 1991).
Содержание плазмина и плазминогена в молоке коров монбельярской породы в течение лактации повышалось с 0,25 до 0,38 и с 1,07 до 2,01 мкг/мл соответственно. Отмечают, что содержание продуктов распада в молоке β-казеина (γ-казеины) к концу лактации повышалось с 3 до 10% (Fox Р. F., 1981; Btnslimane S., Dognin-Bergeret M. J., Berdague J. L. et al., 1990). На рис. 25 показано изменение активности плазмина и плазминогена в сборном сыром молоке коров джерсейской породы в течение лактации.
Также установлено, что активность протеиназы 1 в молоке голштинской породы коров (число бактериальных клеток менее 300 КОЕ тыс/мл) в начале лактации была несколько меньше, чем в более поздний период лактации (через 7...8 мес), и составляла 4,2 и 5,4 ед., что соответствует 0,08 и 0,11 мкг плазмина/мл. В этот же период содержание плазминогена повысилось с 0,5 до 0,9 мкг/мл. Это обусловлено тем, что поздняя стадия лактации коров характеризуется повышенной проницаемостью кровеносных сосудов в молочной железе, сопровождающейся повышенной секрецией различных кофакторов и некоторых ферментов, в том числе и протеиназы I, в молоко (Korycka-Dahl M., Ribadeau Dumas В., Ghene N., 1983). С увеличением числа лактации активность плазмина в молоке коров повышается.
Некоторое различие в содержании ферментов в молоке отмечается в зависимости от порций секретируемого молока. Например, содержание лактопероксидазы и амилаз в молоке меньше в последних порциях молока, чем в первых, а содержание каталазы – наоборот.
На активность ферментов молока оказывают влияние и интервалы между доениями. Так, активность липазы в молоке при сокращении интервала с 12 до 8 или 6 ч повышалась соответственно на 56...116%, а при увеличении интервала до 16 и 18 ч снижалась соответственно на 10 и 12 %. В практических условиях необходимо использовать такие интервалы между дойками, которые не способствовали бы возникновению липолиза в молоке (Suchren G., Hamann J. et al., 1981).
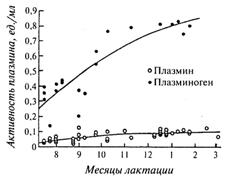
Рис. 25. Изменение активности плазмина (и плазминогена в пересчете на плазмин после активирования урокиназой) в сыром молоке (Richardson В. С, 1983)
При изучении активности ферментов в молозиве и молоке коров установлено, что активность альдолазы с первых суток после отела к пятым увеличивается в 1,2...2,6 раза. Так, у коров черно-пестрой породы высокая активность этого фермента отмечена на 1; 3 и 6 месяцах лактации, а у айрширской породы коров – на 2; 3 и 6 месяцах лактации. В зависимости от возраста достоверные различия обнаружены между коровами первого отела и коровами 2...3 лактации – на 3; 8; 9 и 10 месяцах, а у коров 5...6 отела – на 2; 4; 5; 7; 9 и 10 месяцах. В молоке коров черно-пестрой породы летнего отела активность альдолазы достоверно выше, чем у коров зимнего отела. С удоями молока активность альдолазы в молоке коров указанных пород связана отрицательной корреляционной связью (Курдуманова О. И., 1987).
Таким образом, при оценке в молоке активности нативных ксантиноксидазы, каталазы, карбоксилэстеразы, липазы, щелочной и кислой фосфатаз, лизоцима, протеаз и др. следует учитывать период лактации и молочную продуктивность коров. Эти два зоотехнических фактора являются преобладающими по степени влияния на активность этих ферментов молока.
Время года. Влияние сезона на активность ферментов молока может проявляться в связи с периодом лактации, уровнем молочной продуктивности и кормлением коров. Из рис. 26 видно, что четкого влияния времени года, например, на активность ксантиноксидазы и лактопероксидазы в молоке коров не выявляется. Активность амилазы заметно повышается в середине года, а в остальные месяцы влияние сезона также не выражено. Активность пероксид-дисмутазы в молоке ниже весной, чем осенью и зимой.
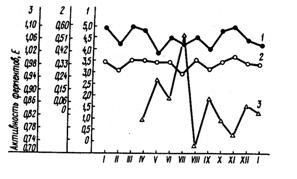
Рис. 26. Сезонные изменения активности ферментов в сыром молоке (по Blanc J., 1982):
1 – лактопероксидаза; 2 – ксантиноксидаза; 3 – амилаза
Поскольку одним из источников аминотрансфераз в молоке является плазма крови, где активность их наиболее высокая летом и осенью, наиболее высокую активность этих ферментов в молоке следует ожидать в это же время года.
В различное время года максимальная и минимальная активности щелочной фосфатазы в сборном сыром молоке отличается примерно в 2 раза (Harding F., 1991). Из рис. 27 видно, что более низкая активность фермента наблюдается летом, а высокая – зимой. Летом активность кислой, а зимой щелочной фосфатазы имеет более широкие пределы колебаний. Так, активность кислой фосфатазы зимой в среднем на 40%, а щелочной – на 10 % выше, чем летом. Зимой отмечается и более высокая активность в молоке холинэстеразы.
Довольно заметное влияние время года оказывает на активность в молоке каталазы и липазы. Это, прежде всего, связано с тем, что осенне-зимний период года в основном совпадает с концом лактации коров, когда активность этих ферментов в молоке, как и протеиназ, заметно увеличивается.
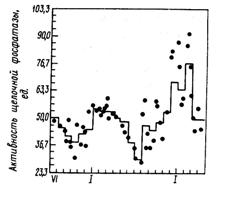
Рис. 27. Сезонные изменения активности щелочной фосфатазы в сыром молоке (1 ед. соответствует образованию фенола мкг/мл/мин при 40 °С) (Murthy G. K., Cox S. et. al.)
Показано, что активность каталазы в свежевыдоенном молоке составляла 8...17% О2 в весенне-летний и 20...25 % О2 в осенний периоды года (Шидловская В. П., Патритий А. П., Насонова Л. М.). На рис. 28 показаны пределы изменения степени липолиза молока в течение года. Максимальные и минимальные значения различаются примерно в 4 раза. Более выраженный липолиз наблюдается осенью. Выявлены следующие закономерности: в июне и марте примерно 22 и 23 % проб молока содержали СЖК более 1,0, около 5 и 8 % - 1,5; 1 и 3 % – 2 мэкв/100 г жира, а в сентябре и декабре примерно 30 и 37 % проб соответственно более 1,0; 10 и 17 % – более 1,5 и 4 и 9 % – более 2,0 СЖК мэкв/100 г жира.
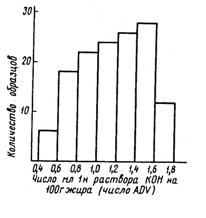
Рис. 28. Содержание свободных жирных кислот в сборном сыром молоке в течение года (по Downey W. К.)
Изучение динамики изменения протеолитической активности молока, на примере черно-пестрой породы коров, показывает, что в молоке коров 3...4 отела в летнее время года до 50 сут после отела она была выше в 3...4 раза, чем в зимнее время года, а в молоке коров айрширской породы – соответственно на 95 % (Ипатова О. М., 1988).
В табл. 23 приведены данные по содержанию плазмина и плазминогена в сборном молоке коров монбельярской породы с 9 ферм и 6-ти сыродельных заводов в зависимости от сезона года (Btnslimane S., . Dognin-Beгgeret М.J., Berdague J. L. et al. 1990).
Таблица 23
 Изучение динамики N-ацетил-β-глюкозаминидазной активности в молоке клинически здоровых 188 коров черно-пестрой породы (содержание соматических клеток не более 500 тыс/мл) при нормальных условиях содержания и кормления (удои за лактацию 4500...5000 л), показало, что несколько повышенный уровень этого фермента встречался с частотой 5...6 %, характеризуя частоту нарушения целостности клеток молочной железы, что может служить показателем, предупреждающим о возможности развития предмаститных состояний или скрыто текущих клинически скрытых маститов (Степанов Г. С., Батраков А. Я. и др., 1988).
Изучение динамики N-ацетил-β-глюкозаминидазной активности в молоке клинически здоровых 188 коров черно-пестрой породы (содержание соматических клеток не более 500 тыс/мл) при нормальных условиях содержания и кормления (удои за лактацию 4500...5000 л), показало, что несколько повышенный уровень этого фермента встречался с частотой 5...6 %, характеризуя частоту нарушения целостности клеток молочной железы, что может служить показателем, предупреждающим о возможности развития предмаститных состояний или скрыто текущих клинически скрытых маститов (Степанов Г. С., Батраков А. Я. и др., 1988).
Таким образом, из приведенных данных видно, что время года наиболее отражается на активности тех ферментов, величина активности которых в молоке наиболее тесно связана с изменением обмена веществ в организме при снижении молочной продуктивности коров.
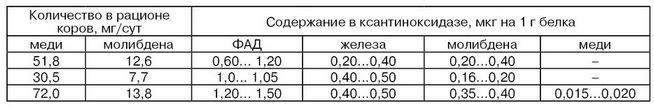
Кормление коров. Фактор кормления коров является одним из главных факторов, влияющих на активность ферментов в молоке. Особый интерес представляет вопрос зависимости активности ферментов молока от содержания микроэлементов в кормовом рационе, так как микроэлементы входят в состав активных центров или выполняют роль активаторов ферментов. На примере ксантиноксидазы (табл. 24) видно, что содержание меди и молибдена в рационе коров заметно влияет на их содержание в молекуле фермента, а следовательно, и на его активность (Ковальский В. В. и др.).
Таблица 24
Кормовой рацион влияет на важнейший фермент антибактериальной активности молока – лактопероксидазу. Так, при скармливании коровам айрширской породы ячменно-овсянной смеси, содержащей ксилит (26,5 %), или мелассового фуража в течение 12 дней содержание этого фермента в молоке в случае использования ксилита составило 17,8 мкг/мл, а в случае мелассы – 12,6 мкг/мл, по сравнению с контролем – 11,7 мкг/мл (Korhonen H., Rintamäki O., Antila M. et а1., 1978).
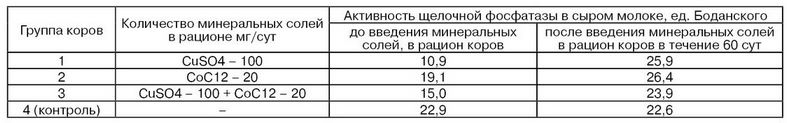
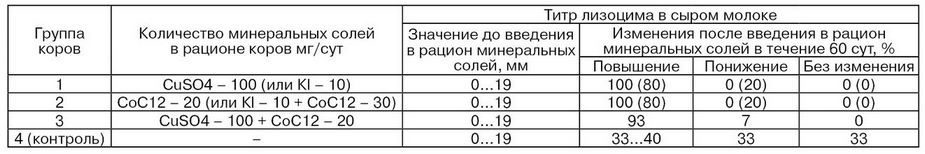
Активность щелочной фосфатазы в молоке заметно снижается при пастьбе коров на пастбище с моховым покровом, содержание марганца в котором примерно на 40 % ниже, чем в растительности на холмистой местности. На рис. 29 показана зависимость между содержанием марганца в сене и молоке и активностью щелочной фосфатазы в молоке. Результаты исследований, проведенных А. С. Авакяном, показывают, что при подкормке коров сульфатом меди и хлоридом кобальта активность щелочной фосфатазы в молоке повышается в 1,5...2,5 раза (табл. 25). Этим же исследователем выявлена прямая зависимость между содержанием в рационе коров микроэлементов и повышением в молоке активности лизоцима (табл. 26). Так, включение в кормовой рацион микроэлементов позволяет значительно повысить низкую активность лизоцима молока – важнейшего фактора неспецифического иммунитета.
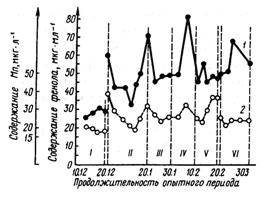
Рис. 29. Зависимость между содержанием марганца и активностью щелочной фосфатазы в сыром молоке (по Тёпел А., 1979):
1 – содержание фенола, мкг/мл; 2 – содержание марганца, мкг/л;
I – сено, обедненное марганцем; II – нормальный период; III – сено, обедненное марганцем; IV – добавлено 3,6 г MnSO4; V – добавлено 3,6 г MnSO4; VI – сено, обогащенное марганцем
Таблица 25
Широкая распространенность спонтанного липолиза на последней стадии лактации коров может быть связана с отсутствием пастбищного кормления или с изменением рациона.
Качество и количество корма являются важными факторами, которые влияют на возникновение спонтанного липолиза в молоке. Низкая энергетическая ценность рациона и высокое содержание пальмитиновой кислоты в нем способствуют развитию в молоке такого липолиза. В период повышенной «чувствительности» молока к липолизу – в конце лактации – влияние качества кормового рациона на липолиз в молоке более выражено. Спонтанный липолиз в молоке связывают с определенными видами кормов, таких, как свекла, сорговой силос, сено плохого качества. Скармливание коровам зеленого корма предотвращает случаи продуцирования молока с последующим развитием в нем липолиза. Так, например, при пастьбе коров на пастбище с молодой травой, для которой характерно высокое содержание ненасыщенных жирных кислот с длинной углеродной цепью (олеиновая, линолевая) и низкое содержание насыщенных жирных кислот со средней углеродной цепью (например, пальмитиновая), в получаемом молоке в значительно меньшей степени возникает спонтанный липолиз. При регулярном скармливании коровам концентратов в молоке также снижается возможность возникновения спонтанного липолиза, при этом содержание в молоке СЖК снижается в 1,5...2,0 и более раз (Chilliard Y., 1982).
Таблица 26
В то же время неудовлетворительный минеральный состав кормов служит причиной спонтанного липолиза молока. Опыты по кормлению коров показывают, что переход с низкого энергетического уровня рациона на высокий уровень приводит к уменьшению возможности возникновения спонтанного липолиза в молоке примерно через 4...5 сут.
Отмечается, что зеленые корма повышают общую оксидазную активность молока в 2...3 раза. В то же время на активность каталазы молока скармливание животным зеленого корма не влияет, тогда как введение в рацион сахарной свеклы повышает ее активность.
Неправильное использование допущенных к применению в сельском хозяйстве химических средств для борьбы с вредителями (например, пестицидов) приводит к попаданию их в корм и воду, а затем в молоко. Имеются некоторые сведения о влиянии на ферменты молока остаточных количеств фосфор- и хлорорганических соединений. Согласно этим данным, одни фосфорорганические пестициды (аминопаратион, хлороксон, карбофос, меркаптофос) заметно подавляют, например, активность лактопероксидазы, обнаруживаемой в сборном молоке. В то же время другие фосфорорганические пестициды (параксон, диизопропилпаратион, диазинон, октаметил, хлорофос), так же как и хлорорганические пестициды (ДДТ, линдан, дилдрин, токсафен, гептахлорэпоксид), не влияют на активность лактопероксидазы даже в значительно большей их концентрации –десятки и сотни миллиграмм на 1 кг. Длительное выдерживание молока, например, с тиофосом при концентрации порядка сотен миллиграмм на 1 кг не вызывало восстановление активности лактопероксидазы, что свидетельствовало о необратимости процесса ингибирования фермента (И. Р. Давидова).
При скармливании животным плесневелых кормов в молоко могут попасть микотоксины, среди которых афлатоксины обладают наибольшей токсичностью. Источниками афлатоксинов в молоке могут быть также соломенная подстилка, навоз и воздух. Афлатоксины начинают выделяться с молоком через 12 ч после скармливания животным инфицированных кормов, и их выделение прекращается спустя 4...5 дней. Эти микотоксины в определенных концентрациях изменяют ферментативную активность молока. Так, наличие афлатоксинов в молоке может заметно (в 3...4 раза) повысить активность кислой фосфатазы. Несмотря на то, что активность этого фермента в молоке невелика (до 10% активности щелочной фосфатазы), он обладает очень высокой термоустойчивостью, и проявление его активности в питьевом молоке нежелательно, так как способствует снижению его качества.
[1] 1 ед. активности – см. сноску на с 77.


